RESYNTHESE DE
L’ATP – LE METABOLISME ENERGETIQUE :
I – PRESENTATION GLOBALE DU METABOLISME ENERGETIQUE :
1-1 :
INTRODUCTION :
Il se passe à l’intérieur de toutes les cellules. Il a pour fonction de resynthétiser l’ATP (adénosine triphosphate), grâce à la dégradation de substrats énergétiques. Ces substrats circulent dans le sang et proviennent essentiellement de la dégradation de la digestion. Quand ils sont dans le sang, ils sont captés par les cellules selon les besoins.
Certains types cellulaires ont la possibilité de bloquer ces substrats. C’est le cas de la cellule musculaire, que l’on appelle aussi des fibres musculaires, qui stockent le glucose sou forme de glycogène. De même les acides gras sont stockés par les cellules graisseuses sous forme de triglycérides. Ces stockes existent de pour qu’ils soient utilisés sous forme de substrats énergétiques. Les cellules du foie, les hépatocytes, peuvent aussi stocker le glucose sous forme de glycogène. Ce glucose hépatique peut recirculer dans le sang pour être capté par d’autres types cellulaires, tels que les cellules musculaires. Le foie est donc capable de stocker les substrats et parfois de les redistribuer. Tel est le cas des aussi des adipocytes (cellules graisseuses), qui vont stocker les acides gras sous forme de triglycérides. Ces substrats énergétiques vont être dégradés au sein de la cellule grâce à des réactions biochimiques et qui sont catalysés par des enzymes. Certaines de ces réactions sont appelées exergoniques et libèrent de l’énergie, dont une partie va permettre de resynthétiser l’ATP, c’est-à-dire de créer au sein de cette molécule des liaisons riches en énergie.
Le métabolisme énergétique est composé de différents systèmes, qui sont composés d’une suite de réactions chimiques qui s’enchaînent les unes aux autres. On les appelle le système polyenzymatique. Une partie de ces réactions est spécifique à un substrat et les autres sont communes aux autres substrats. On note plusieurs exemples de systèmes spécifiques :
· La glycolyse, qui est spécifique au glucose.
· La béta-oxydation, qui est spécifique à la dégradation des acides gras.
On trouve aussi des exemples de systèmes non-spécifiques :
· Le cycle de Krebs
· La chaîne respiratoire.
Au point de vue bioénergétique, c’est la chaîne respiratoire, qui est le lieu principal de resynthèse de l’ATP. Elle se passe au niveau des mitochondries, que l’on trouve notamment dans les cellules musculaires. Cette chaîne respiratoire ne peut fonctionner qu’à l’aide des autres systèmes polyenzymatiques. En effet des derniers produisent les substrats de la chaîne respiratoire.
Il existe donc deux substrats énergétiques, qui donnent deux métabolismes : Le métabolisme glucidique et le métabolisme lipidique. Tous les deux convergent vers le cycle de Krebs, qui est suivi par la chaîne respiratoire, qui est aussi commune aux deux métabolismes.
Enfin le glucose provient soit du sang soit du glycogène. Les acides gras proviennent soit du sang, soit des triglycérides.
1-2 :
DEFINITIONS BASIQUES :
1-2-1 :
Métabolisme énergétique :
C’est un ensemble de systèmes polyenzymatiques, qui a pour fonction de resynthétiser l’ATP grâce à la dégradation des substrats énergétiques. On peut appeler cette dégradation le catabolisme. Il comporte aussi des systèmes polyenzymatiques, qui stockent et déstockent des substrats énergétiques.
Comme les substrats énergétiques les plus fréquents sont le glucose et les acides gras, le métabolisme énergétique comporte deux systèmes :
· Le système glucidique.
· Le système lipidique.
1-2-2 :
L’ATP :
C’est une molécule qui « stocke » l’énergie sous forme chimique et en particulier sous forme de liaisons chimiques avec du phosphate. C’est la seule molécule de l’organisme qui peut restituer de l’énergie pour faire fonctionner les réactions qui en ont besoin, c’est-à-dire les réactions endergoniques (qui ont besoin d’énergie).
1-2-3 :
Substrats énergétiques :
Ce sont des composés biochimiques, qui une fois dégradés permettent de resynthétiser de l’ATP. Les principaux substrats énergétiques sont les glucides et les lipides. Ils peuvent exister sous forme libre ou sous forme de stockage (glycogène et triglycéride). Le glycogène est un polymère du glucose et les triglycérides correspondent à trois acides gras et à un glycérol.
Le glucose provient soit de l’alimentation, soit d’une synthèse réalisée par l’organisme à partir de précurseurs du glucose. La synthèse de glucose à partir de précurseurs s’appelle néoglucogenèse.
Les acides gras, stockés dans les muscles et les adipocytes, provient de l’alimentation ou d’une synthèse à partir de précurseurs.
1-2-4 :
Le métabolisme glucidique :
Il est notamment composé de systèmes polyenzymatiques, qui concernent le stockage sous forme de :
· Glycogenèse, c’est-à-dire la synthèse du glycogène.
· Glycogénolyse, c’est-à-dire la destruction de glycogène.
Les autres systèmes sont :
· La synthèse de glucose à partir de précurseurs (néoglucogenèse).
· Le début de la dégradation du glucose (glycolyse).
· Le cycle de Krebs.
· La chaîne respiratoire.
1-2-5 :
La glycogénolyse :
C’est un système polyenzymatique, qui a pour substrat le glycogène et pour produit le glucose. Le glucose est donc déstocké. Seules les cellules musculaires et hépatiques peuvent faire de la glycogénolyse.
1-2-6 :
La glycogenèse :
C’est un système polyenzymatique, dont le substrat énergétique est le glucose et dont le produit est le glycogène. Le glucose est ici stocké et seules les cellules musculaires et hépatiques ont la capacité à réaliser la glycogenèse.
1-2-7 :
La néoglucogenèse :
C’est un système polyenzymatique, dont le substrat énergétique est un précurseur du glucose, tel que le lactate ou les acides gras et dont le produit est le glucose. On refait donc ici de nouvelles molécules de glucose et là encore seules les cellules musculaires et hépatiques sont aptes à réaliser la néoglucogenèse.
1-2-8 :
La glycolyse :
C’est un système polyenzymatique, dont le substrat énergétique est le glucose et dont le produit est le pyruvate et le lactate dans quelques occasions. Sur le plan énergétique, il permet la resynthèse d’une petite quantité d’ATP et de NADH.
Toutes les réactions citées auparavant (glycogénolyse, glycogenèse, néoglucogenèse, glycolyse) sont donc en rapport avec le métabolisme glucidique.


1-2-9 : Le métabolisme
énergétique des acides gras :
Il est composé de systèmes polyenzymatiques qui concernent le stockage des acides gras sous forme de triglycérides, le déstockage des acides gras à partir de triglycérides, la synthèse d’acides gras à partir de précurseurs et la dégradation des acides gras.
1-2-10 : La
lipolyse :
C’est un système polyenzymatique, qui a pour substrat les triglycérides et produit les acides gras + le glycérol. Les acides gras sont donc déstockés. Les adipocytes, c’est-à-dire les cellules graisseuses, et les cellules musculaires sont capables de lipolyse.
1-2-11 : La
lipogenèse :
C’est un système polyenzymatique dont le substrat énergétique est un acide gras et un glycérol et dont le produit est un triglycéride. C’est la forme de stockage des acides gras et ce système ne se produit que dans les adipocytes et dans les cellules musculaires.
1-2-12 : La
béta-oxydation :
C’est un système polyenzymatique, dont le substrat est un acide gras et dont le produit est l’acétyl-CoA.
La béta-oxydation ne resynthétise pas directement de l’ATP mais produit du NADH et du FADH.

1-2-13 : Le cycle de
Krebs :
On l’appelle aussi cycle cytrique et cycle tricarboxylique.
C’est un cycle qui est alimenté par l’acétyl-CoA et dont le produit est le CO2 qui est expiré. C’est donc un système polyenzymatique qui resynthétise une petite quantité d’ATP et produit du NADH et du FADH.
L’acétyl-CoA peut avoir comme origine le glucose ou la béta-oxydation et est par conséquent produit par les deux types de métabolismes énergétiques. C’est la suite commune de la dégradation complète (produit : CO2 + H2O) du glucose et des acides gras.
1-2-14 : NAD et
FAD :
Ce sont des co-enzymes, appelées aussi des déshydrogénases. Ils captent les électrons arrachés aux substrats lors des réactions d’oxydo-réductions qui ont lieu au niveau de la glycolyse, de la béta-oxydation et du cycle de Krebs.
Après avoir capté ces électrons, NAD est réduit en NADH et FAD est réduit en FADH. FADH et NADH vont ensuite transporter leurs électrons jusqu’à la chaîne respiratoire. On les appelle aussi des transporteurs d’électrons :
![]() NAD + 2 H NADH2 = NADH,H.
NAD + 2 H NADH2 = NADH,H.
1-2-15: La chaîne
respiratoire:
On parle de phosphorilation oxydative.
C’est un système polyenzymatique, dont le substrat est NADH, FADH etO2 et dont le produit est H2O et de l’ATP en grande quantité.
Les électrons de NADH et de FADH sont transportés par des transporteurs de la chaîne respiratoire jusqu’à l’oxygène qui est le dernier accepteur de l’électron et qui va être réduit en eau.
L’oxydation de NADH et de FADH produit de l’énergie, qui va permettre de resynthétiser de l’ATP en grande quantité et ceci grâce à l’enzyme : ATP synthétase. La chaîne respiratoire est le point final de la dégradation des substrats.
1-3 :
REACTION ANDERGONIQUE ET EXERGONIQUE :
Lorsqu’un composé chimique est entièrement brûlé, il libère de la chaleur qu’on appelle anthapie. Quand un substrat énergétique est entièrement oxydé, il libère de l’énergie. Seule une partie de cette énergie sera utilisable pour l’organisme.
1-3-1 : Dégradation du
glucose :
La dégradation complète du glucose suit l’équation bilan suivante :
![]() C6H12O6
+ 6O2
6H2O + 6CO2.
C6H12O6
+ 6O2
6H2O + 6CO2.
Cette réaction va libérer de l’énergie, mais seule une partie sera utilisée.
1-3-2 : Dégradation des
acides gras :
La dégradation complète des acides gras suit l’équation bilan suivante :
![]() Acide
gras + xO2
yH2O + zCO2.
Acide
gras + xO2
yH2O + zCO2.
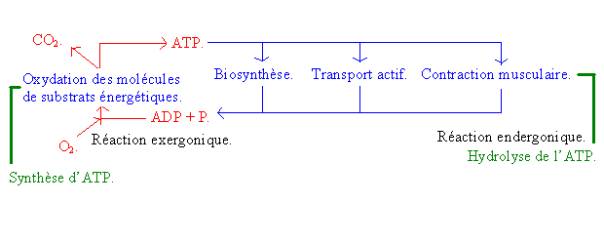
Les réactions qui libéreront de l’énergie seront appelées réactions exergoniques et les réactions qui utilisent de l’énergie seront appelées andergoniques. Ainsi les réactions exergoniques permettent aux réactions andergoniques d’avoir lieu.
Parmi les réactions, une nous intéresse particulièrement : Il s’agit de celle qui amène à la synthèse le l’ATP selon l’équation bilan :
![]() ADP + P ATP.
ADP + P ATP.
On trouve aussi parmi les réactions andergoniques :
· La contraction musculaire, qui est provoquée par une enzyme qui active la myosine et qui consomme de l’énergie.
· Tous les transports : ions : calcium (dans le muscle il s’agit du calcium ATPase qui pompe le calcium de la myofibrille).
Donc il existe un couplage entre les deux types de réactions qui est assuré par l’ATP.
1-4 :
L’ATP :
C’est une molécule qui stocke de l’énergie sous forme d’énergie chimique. C’est quasiment la seule qui peut restituer l’énergie nécessaire aux réactions andergoniques. L’ATP stocke cette énergie grâce à ces liaisons avec les groupements phosphates. Celles-ci sont dites riches en énergie si leur rupture libère plus de 5 kcal ( = 21 kJ).
L’ATP comporte essentiellement deux liaisons riches en énergie. Elles peuvent être cassées par hydrolyse :
· Première réaction :
![]()
![]() ATP
+ H2O
ADP + PO4H3 : Cette réaction est ainsi
déséquilibrée et incomplète. On va la simplifier et noter : ATP ADP + P.
ATP
+ H2O
ADP + PO4H3 : Cette réaction est ainsi
déséquilibrée et incomplète. On va la simplifier et noter : ATP ADP + P.
Il s’agit de la réaction qui casse donc la première liaison riche.
· Deuxième réaction :
![]() ADP
+ H2O AMP +
PO4H3 : Là encore on note en simplifiant :
ADP
+ H2O AMP +
PO4H3 : Là encore on note en simplifiant :
![]() ADP AMP + P, avec
AMP : Adénosine monophosphate.
ADP AMP + P, avec
AMP : Adénosine monophosphate.
En général, le dernier phosphate reste accroché.
![]() La
phosphocréatine est de même que l’ATP capable de stocker l’ATP selon l’équation
bilan : CP + ADP
C + ATP, avec C : la créatine.
La
phosphocréatine est de même que l’ATP capable de stocker l’ATP selon l’équation
bilan : CP + ADP
C + ATP, avec C : la créatine.
La créatine phosphate a elle une liaison riche en énergie.
1-5 : LES
REACTIONS D’OXYDO-REDUCTIONS :
Ce sont des réactions biochimiques, catalysées par des déshydrogénases (ou oxydases ou encore réductases). Lors d’une réaction d’oxydation, le substrat, qui est oxydé, perd son électron. De même lors d’une réaction de réduction, le substrat, qui est réduit, capte un électron. Les réactions sont la plus part du temps couplées entre elles.
![]()
![]() Composé
réduit (oxydation) composé oxydé.
Composé
réduit (oxydation) composé oxydé.
![]()
![]() Composé
oxydé (réduction) composé réduit.
Composé
oxydé (réduction) composé réduit.
![]()
![]() A
réduit + B oxydé (oxydation
de A ; réduction de B) A
oxydé + B réduit.
A
réduit + B oxydé (oxydation
de A ; réduction de B) A
oxydé + B réduit.
Dans la plus part des cas, dans le métabolisme énergétique, le composé qui est réduit est NAD pour devenir NADH ou FAD pour devenir FADH. Ce sont les transporteurs d’électrons qui sont réduits et se sont les co-enzymes des déhydrogénases.

II – STOCKAGE ET DESTOCKAGE :
2-1:
INTRODUCTION:
Dans une situation où il n’y a pas d’apport alimentaire, les substrats lipidiques ou glucidiques sont déstockés afin de permettre la resynthèse de l’ATP. Après un effort ou un moment de privation d’alimentation, les stocks de glycogènes et de triglycérides ont besoins d’être restitués. C’est fait grâce à l’apport alimentaire.
2-2 : LA
GLYCOGENESE (SYNTHESE DE GLYCOGENE A PARTIR DE GLUCOSE) :
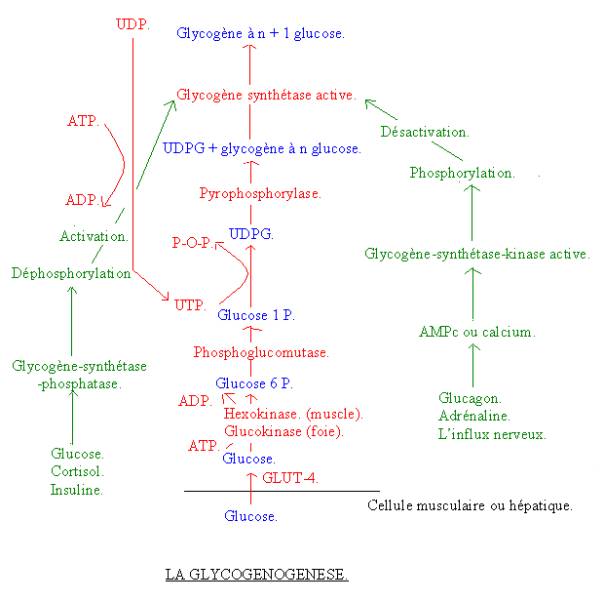
La glycogenèse ne se produit que dans les cellules musculaires ou les cellules hépatiques.
2-2-1 : Les différentes
réactions :
· Le glucose entre dans la cellule musculaire grâce à un transport : Le transporteur GLUT-4, qui est le transporteur du glucose pour la fibre musculaire. C’est une protéine de la membrane qui fait entrer le glucose dans la cellule.
· La phosphorylation : Elle consiste à transformer le glucose, qui est alors le substrat en glucose 6 P, qui est lui le produit. La réaction est catalysée par l’hexokinase. C’est une réaction de phosphorylation du glucose, qui consomme une ATP, qui s’hydrolyse en ADP.
Cette réaction suit l’équation bilan suivante :
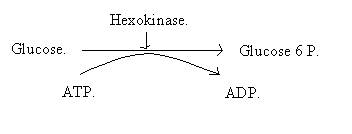
· Le glucose 6 P se transforme en glucose 1 P. Le phosphate passe donc du carbone n°6 sur le carbone n°1. Cette réaction est catalysée par une phosphoglucomutase.
· Le glucose 1 P auquel vient s’ajouter un UTP se transforme en UDPG, l’UTP étant une molécule très proche de l’ATP. Cette réaction consomme un second ATP sous forme d’UTP. Cette réaction est catalysée par la UDPG pyruphosphorilase.
· L’UDPG auquel vient s’ajouter une molécule de Glycogène à n molécule de glucose donne un glycogène à n + 1 molécule de glucose. Le polymère du glucose qui existe s’enrichit d’une molécule de glucose. Cette réaction est catalysée par la glycogène synthétase. Cette enzyme ne va catalyser cette réaction que si elle se trouve sous forme active.
Tout ça ne se produit que le foie et le muscle.
2-2-2 : La régulation
de la glycogénogenèse :
La régulation va se faire au niveau du passage de la forme active ou inactive de la glycogène-synthétase. Cette enzyme est transformer elle-même par deux autres enzymes, qui sont elles-même déclenchées par un système et des hormones (insuline, glucagon, …). Par conséquent :
· L’adrénaline, le glucagon et l’influx nerveux inhibent la glycogénogenèse. Ils vont en effet produire un AMP cyclique (AMPc), qui est un second messager ou du calcium. Ces deux derniers vont activer une enzyme : la glycogène synthétase kinase. Quand celle-ci est active, elle transforme la forme active de la glycogène synthétase en forme inactive. Il y a alors inhibition de la glycogénogenèse.
· Le glucose (substrat énergétique), le cortisol (hormone produite par la glande corticosurrénale) et l’insuline (hormones produites par le pancréas) activent la glycogénogenèse. Elles vont en effet activer l’enzyme glycogène synthétase phosphatase, qui transforme la forme inactive glycogène synthétase en forme active. La glycogénogenèse peut alors se faire.
2-3 : LA
LIPOGENESE : SYNTHESE DE TRIGLYCERIDES :
2-3-1 : Les différentes
réactions :
· Trois acides gras et trois co-enzymes A et trois ATP vont donner trois acyl CoA. La réaction va être catalysée par l’acide gras thiokinase.
· Deux acyl CoA vont se combiner avec le glycérol pour former un diglycéryl et deux 2 CoA. La réaction va être catalysée par la phosphotide synthétase et phosphatase.
· On rajoute un troisième acyl CoA pour former un triglycéride et un CoA. La réaction est ici catalysée par la triglycéride synthétase.
2-3-2 : La
régulation :
· L’insuline active la lipogenèse.
· Les catécholamines inhibent la lipogenèse. Elles sont produites par la glande médullosurrénale.
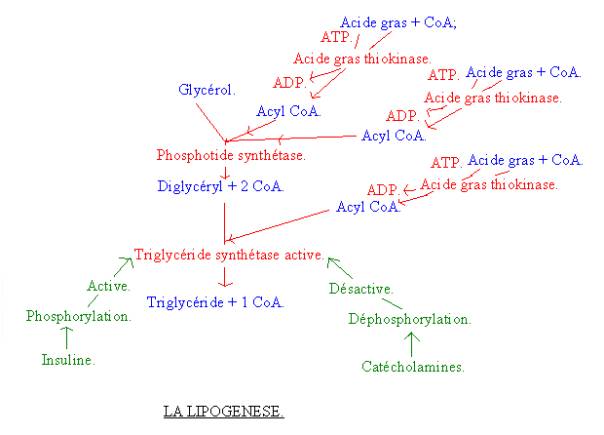
La régulation donc se fait là-encore par des hormones.
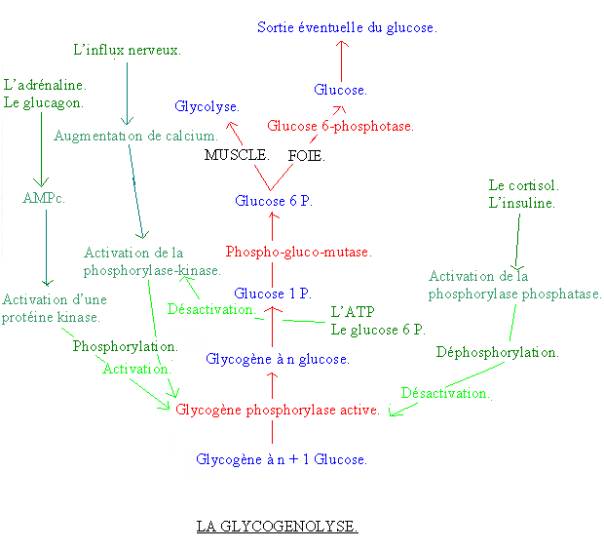
2-4 : LA GLYCOGENOLYSE :
2-4-1 : Les différentes
réactions :
· Une molécule de glucose 1 P se sépare de la molécule de glycogène. Cette réaction est catalysée par la glycogène phosphorylase, si elle se trouve sous forme active.
· Le glucose 1 P devient alors du glucose 6 P. Cette réaction est catalysée par la phosphoglucomutase.
· Le glucose 6 P devient du glucose. Ceci est réalisé que dans le foie par l’enzyme glucose 6 phosphatase. Ce glucose hépatique peut éventuellement sortir du foie pour être véhiculé dans le sang vers les organes qui en ont besoin.
Dans le muscle, le glucose 6 P est transformer par la glycolyse.
2-4-2 : La
régulation :
La régulation se fait au niveau de la glycogène phosphorylase, qui existe sous forme active ou inactive.
· L’influx nerveux active la glycogénolyse via une augmentation de calcium. Cette augmentation de calcium va activer la phosphorilase-kinase, qui va elle-même transformer la forme inactive de la glycogène phosphatase en forme active.
· L’adrénaline et le glucagon activent également la glycogénolyse. Il y a une production d’AMPc, qui active une protéine kinase. Celle-ci va alors transformer la forme inactive de la glycogène phosphorylase kinase en forme active.
· L’ATP et le glucose 6 P inhibent la glycogénolyse en inhibant la glycogène-phosphatase kinase.
· Le cortisol et l’insuline inhibent aussi la glycogénolyse en activant la phosphorylase-phosphatase, qui se transforme alors la forme active de la glycogène-phosphorylase en forme inactive.
2-5 : LA LIPOLYSE :
On va passer du triglycéride aux acides gras.
2-5-1 : Les différentes
réactions :
· Le triglycéride se scinde en un diglycéride et en un acide gras grâce à une enzyme appelée la triglycéride-lipase, quand elle est sous forme active.
· Le diglycéride se scinde lui aussi en un monoglycéride et en un acide gras, grâce à la diglycéride-lipase.
· Le monoglycéride se scinde en un acide gras et en un glycérol. La réaction est catalysée par la monoglycéride-lipase.
Si l’acide gras se trouve dans un adipocyte, il peut alors en sortir pour aller dans le sang et être capter par d’autres cellules. Par contre si l’acide gras est dans le muscle, il y reste et est dégradé par la béta-oxydation.
2-5-2 : La
régulation :
Elle se fait au niveau de la triglycéride-lipase, qui existe sous forme active ou inactive. Ainsi :
· L’insuline inhibe la lipolyse. Elle active une phosphoprotéine, qui transforme la forme active de la triglycéride-lipase en forme inactive.
· L’adrénaline et le glucagon active la lipolyse. Elles conduisent en effet à la formation d’AMPc qui active une protéine kinase, qui transforme la forme inactive de la triglycéride-lipase en forme active.
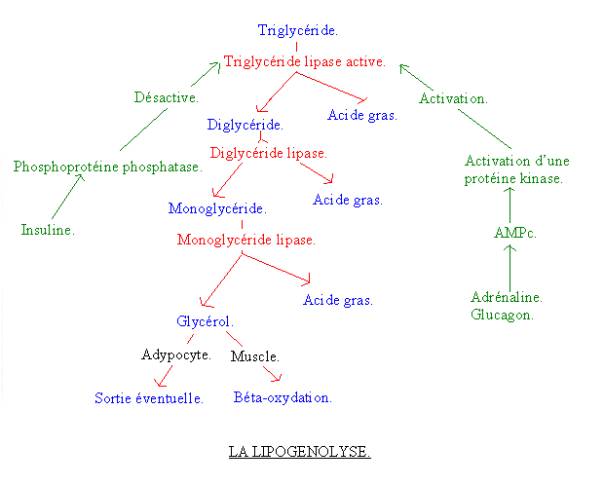
Finalement on a à faire à une substance qui va inhiber le déstockage du substrat et activer son stockage et inversement. De plus l’insuline a généralement l’effet inverse au catécholamine. Il est également normal que l’influx nerveux active le déstockage du substrat car ces substrats pourront être dégradés pour former de l’ATP.
|
|
Glycogénogenèse. |
Lipogenèse. |
Glycogénolyse. |
Lipolyse. |
|
Ca2+ |
- |
|
+ |
|
|
Adrénaline. |
- |
- |
+ |
+ |
|
Insuline. |
+ |
+ |
- |
- |
|
Cortisol. |
+ |
|
- |
|
III – LA
GLYCOLYSE :
La glycolyse est une suite de réactions enzymatiques et est la voie de dégradation des oses composés de 6 carbones (hexoses, glucose). La glycolyse a pour but de dégrader une molécule à 6 carbones pour en faire deux molécules à 3 carbones, c’est-à-dire deux pyruvates ou deux lactates. Pour cela elle comporte une dizaine de réactions d’états biochimiques qui s’enchaînent. Certaines réactions sont irréversibles.
Tout d’abord il faut que le glucose entre dan la cellule, grâce à un transporteur le GLUT-4 ( = glucose transporteur n°4).
3-1 : LA
PHOSPHORILATION DU GLUCOSE :
Durant cette première phase, le glucose va être transformer en glucose 6 P. Cette réaction nécessite aussi la dégradation D’une molécule d’ATP en ADP. Cette réaction est irréversible. Le glucose est donc phosphorylé. Le phosphate que perd l’ATP va en effet venir sur le sixième carbone du glucose. La réaction va donc consommer un ATP et est catalyser par une hexokinase (la glucokinase).
![]() Glucose + ATP Glucose 6 P +
ADP.
Glucose + ATP Glucose 6 P +
ADP.
3-2: FORMATION DU
FRUCOSE 6 P :
Cette réaction est une isomérisation et est réversible. Elle est catalysée par la phosphoglucose isomérase et permet la transformation du glucose 6 P en fructose 6 P.
3-3 :
FORMATION DU 1,6 DIP :
C’est une réaction de phosphorylation qui est irréversible et catalysée par la phosphofructo kinase (PFK). Cette PFK est une enzyme allostérique, qui est très importante pour la régulation. Cette réaction permet donc la transformation du fructose 6 P en fructose 1,6 DiP, ce qui nécessite la dégradation d’une molécule d’ATP en ADP.
![]() Fructose 6 P + ATP Fructose 1,6 DiP + ADP.
Fructose 6 P + ATP Fructose 1,6 DiP + ADP.
3-4: RUPTURE DU
FRUCTOSE 1,6 DIP:
On va passer d’une molécule à 6 carbones à deux molécules à trois carbones : Le dihydroxyacétone phosphate et le glycéraldéhyde 3-phosphate. C’est une réaction irréversible et catalysée par l’aldolase. Seul le glyceraldehyde 3-phosphate continue la glycolyse.
3-5 :
TRANSFORMATION DU DIHYDROXYACETONE PHOSPHATE :
C’est une réaction irréversible catalysée par la triose phosphate isomérase. Elle permet le passage du dihydroxyacétone phosphate en glycéraldéhyde 3-phosphate.
3-6 :
OXYDATION DU GLYCERALDEHYDE 3-P :
Le glycéraldéhyde 3 P est transformé en 1,3 diphosphoglycérate. Cette réaction est irréversible et est catalysée par la glycéraldéhyde 3-phosphate déshydrogénase. En même temps, il y a une réduction de NAD en NADH,H+ (première réaction de la glycolyse où NAD est réduit). Dans la molécule de 1,3 diphosphoglycérate est crée une liaison riche en énergie.
3-7 :
DEPHOSPHORYLATION DE 1,3 DIPHOSPHOGLYCERATE :
C’est une réaction réversible et catalysée par la phosphoglycérate kinase. Une molécule d’ATP est resynthétisée. L’énergie, qui permet cette resynthèse, vient du fait que la liaison riche en énergie est rompue. Cette réaction va former le 3-phosphoglycérate.
![]()
![]() 1,3
diphosphoglycérate + ADP ATP + 3-phosphoglycérate.
1,3
diphosphoglycérate + ADP ATP + 3-phosphoglycérate.
3-8 :
FORMATION DU 2-PHOSPHOGLYCERATE :
C’est une réaction catalysée par la phosphoglycéromutase et réversible. Le phosphate du 3-phosphoglycérate est tranférer du troisième carbone sur le deuxième carbone, afin de former du 2-phosphoglycérate.
3-9 :
FORMATION DU PHOSPHOENOLPYRUVATE :
![]() C’est
une réaction réversible catalysée par l’énolase. Il y a réorganisement
moléculaire du substrat avec la perte d’une molécule d’H2O. Il y a
une création d’une deuxième liaison riche en énergie au sein du
phosphoenolpyruvate.
C’est
une réaction réversible catalysée par l’énolase. Il y a réorganisement
moléculaire du substrat avec la perte d’une molécule d’H2O. Il y a
une création d’une deuxième liaison riche en énergie au sein du
phosphoenolpyruvate.
![]() 2-phosphoenolpyruvate
phosphoenolpyruvate + H2O.
2-phosphoenolpyruvate
phosphoenolpyruvate + H2O.
3-10:
DEPHOSPHORYLATION DU PHOSPHOENOLPYRUVATE:
C’est une réaction irréversible catalysée par une pyruvate kinase. Il y a alors resynthèse d’ATP à partir d’une molécule d’ADP. Cette réaction permet de créer du pyruvate.
![]() Phosphoenolpyruvate + ADP ATP +
pyruvate.
Phosphoenolpyruvate + ADP ATP +
pyruvate.
L’énergie provient de la rupture de liaison riche en énergie du phosphoenolpyruvate.
Seul le pyruvate sera par la suite dégradé dans le cycle de KREBS. Dans certaines conditions d’exercice, la glycolyse sera plus rapide que le cycle de KREBS. Le pyruvate est réduit en lactate et parallèlement à cela le NADH est oxydé. Cette réaction est réversible et catalysée par la lacticodéshydrogénase.
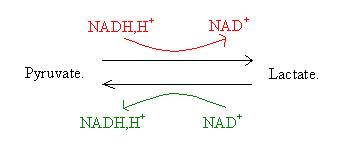
Pour les autres glucides, tels que le glycogène et le galactose sont transformés en glucose 6 P. Ce fructose se transforme en fructose 6 P. A partir delà ils suivent les mêmes réactions que le glucose.
3-11 : BILAN
BIOENERGETIQUE :
Au cours de la glycolyse, il y a donc :
· Synthèse d’ATP :
|
|
ATP. |
|
|
|
Glucose sanguin. |
Glucose du glycogène. |
|
Glucose en glucose 6 P. |
-1 |
0 |
|
Fructose 6 P en fructose 1,6 DiP. |
-1 |
-1 |
|
(2 *) 1,3-Diphosphoglycérate en 3-phosphoglycérate. |
2 * 1 = 2 |
2 * 1 = 2 |
|
(2 *) Phosphoenolglycérate en pyruvate |
2 * 1 = 2 |
2 * 1 = 2 |
|
|
= 2 |
= 3 |
· Synthèse de NADH,H+ :
|
|
NADH produit par glycolyse. |
|
|
|
2 pyruvates. |
2 lactates. |
|
(2 *) glycérate 3 P en 1,3 Diphosphoglycérate. |
2 |
2 |
|
(2 *) pyruvate en lactate. |
0 |
2 * 1 = 2 |
|
|
= 2 |
= 0 |
· On note aussi la présence d’enzymes (4 sur les 11) qui phosphorylent ou qui déphosphorylent (+1 P ou –1 P). Ainsi les isomérases mutases vont être responsables des transformations de configuration.
3-12 : LA
REGULATION :
Les régulations se font à différents niveaux de la glycolyse :
· Le premier niveau se trouve au niveau du GLUT-4, qui est le transporteur du glucose dans la cellule. Ce transport est activé par l’insuline et l’exercice musculaire.
· Le deuxième niveau se fait sur l’hexokinase. S’il y accumulation de glucose 6 P dans les fibres musculaires, l’enzyme est alors inhibée.
· Le troisième niveau de régulation intervient au niveau de la phosphofructokinase, qui est une enzyme allostérique. Elle est activée par l’insuline et par une faible quantité d’ADP. Elle est par contre inhibée par une forte concentration d’ATP ou de NADH, et d’acide citrique, qui est un composé du cycle de KREBS.
· Le quatrième niveau se fait sur la pyruvate kinase. Elle est activée par l’insuline (hormone hypoglmycémiante) et est inhibée par les catécholamines (= adrénaline et le glucagon).
IV – LE CYCLE DE KREBS :
On l’appelle aussi le cycle citrique ou le cycle tricarboxylique. C’est la suite de la dégradation du glucose, qui se produit dans la mitochondrie. Il fait parti du phénomène d’oxydation des substrats, c’est-à-dire qu’il s’agit d’un processus qui nécessite de l’oxygène.
C’est la dégradation de l’acétyl CoA et c’est le point de convergence de la glycolyse et de la bétaoxydation.
4-1 : LE
TRANSPORT DU PYRUVATE EN ACETYL CoA :
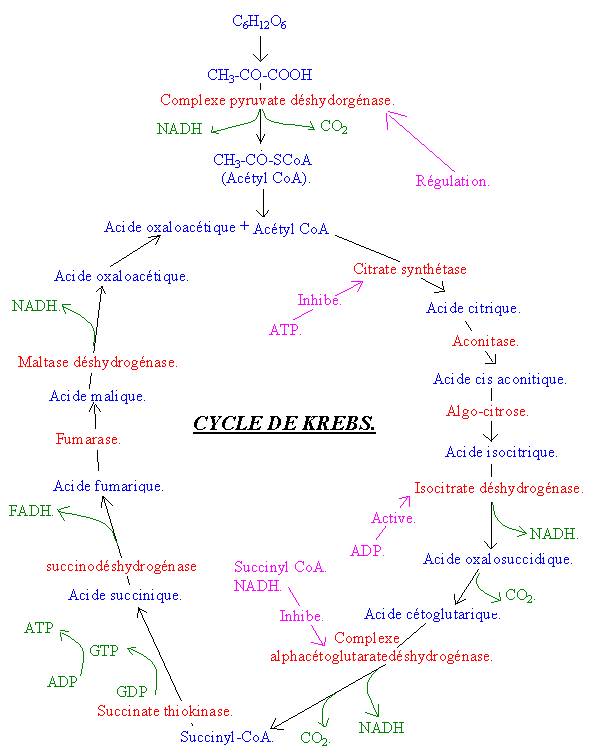
Le pyruvate est une moitié du glucose, puisqu’il n’a que trois carbones. Ce dernier va donner l’acétyl CoA qui possède deux carbones. La réaction est catalysée par le complexe pyruvate déshydrogénase. Cette réaction comprend :
· L’oxydation du pyruvate et la réduction en parallèle du NAD en NADH.
· Une décarboxylation, c’est-à-dire la perte d’un CO2 de la molécule de pyruvate.
C’est une étape très importante dans la régulation de l’oxydation du glucose, c’est-à-dire dans sa dégradation complète.
Cette réaction est inhibée par ces produits, à savoir le NADH et l’acétyl CoA. Aussi ce complexe enzymatique est activé par l’ADP et l’insuline.
![]() Pyruvate + NAD Acétyl CoA + CO2 + NADH.
Pyruvate + NAD Acétyl CoA + CO2 + NADH.
Le pyruvate vient du cytoplasme et entre dans la mitochondrie et la réaction se fait dans la mitochondrie. L’entrée se fait grâce à un transporteur du pyruvate.
4-2 : LE
CYCLE DE KREBS :

4-2-1 : La formation de
l’acide citrique :
L’acétyl CoA et l’acide oxaloacétique vont donner de l’acide citrique. On a donc une molécule à 4 carbones et une molécule à 2 carbones qui donnent une molécule à 6 carbones ; Cette réaction est irréversible et est catalysée par la citrate synthétase, qui est une ligase.
4-2-2 : Formation de
l’acide cis aconitique et d’acide isocitrique :
La formation d’acide cis aconitique à partir l’acide citrique est permis grâce à l’ aconitase. Dans le même temps, il y a formation d’acide isocitrique ou d’isocitrate sous forme ionique. Cette réaction est alors catalysée par l’algocitrase.
4-2-3 : Formation de
l’oxalosuccinate (ou acide oxalosuccidique) :
Il s’agit d’une déhydrogénation, c’est-à-dire une oxylation de l’acide isocitrique. Conjointement il y a réduction de NAD en NADH. Cette réaction est catalysée par l’isocitricodéshydrogénase. On est toujours à une molécule à 6 carbones.
4-2-4 : Formation
d’acide cétoglutarique :
Il s’agit d’une décarboxylation oxydative. Il y a donc départ d’un premier CO2 : On passe alors à une molécule à 5 carbones. Le CO2 ici perdu est alors expiré.
4-2-5 : Formation de
succinyl-CoA :
Cette réaction est composée de deux réactions :
· L’oxydation du cétoglutarate avec réduction de NADH.
· La décarboxylation oxydative, qui correspond au départ d’une deuxième molécule de carbone, qui va être elle aussi expirée. On passe alors à une molécule à 4 carbones. Cette réaction est catalysée par le complexe alphacétoglutaratedéshydrogénase.
4-2-6 : Formation de
l’acide succinique :
Le succinyl CoA et un phosphate et un GDP (molécule proche de l’ADP) donne de l’acide succinique et une molécule de GTP. La réaction est catalysée par la succinate thiokinase. C’est la seule réaction qui permet la resynthèse d’une molécule proche de l’ATP.
Cette réaction libère suffisamment d’énergie pour permettre la création d’une liaison riche en énergie dans la molécule de GDP.
4-2-7 : Formation de
l’acide fumarique :
Il y a oxydation de l’acide succinique et réduction conjointe de FAD en FADH. Cette réaction est catalysée par l’enzyme succinodéshydrogénase.
4-2-8 : Formation
d’acide malique :
L’acide fumarique capte une molécule d’eau. La réaction est catalysée par la fumarase.
4-2-9 : Formation de
l’oxaloacétate (= acide oxaloacétique) :
Il y oxydation de l’acide malique et réduction de NAD en NADH. Le catalyseur est la maltate déshydrogénase (ou malicodéhydrogénase). On finit donc avec une molécule à 4 carbones, ce qui permet de nouveau de réaliser la première réaction.
4-2-10 : Bilan
bioénergétique :
Au total il y a production de 4 NADH, de 1 FADH et de 1 ATP par molécule de pyruvate. Or le glucose produit deux pyruvates. Il faut donc multiplier toutes les quantités par deux quand on parle en molécule de glucose.
4-3 : LES
REGULATIONS :
La régulation se fait à différents niveaux :
· Le premier niveau de régulation se fait au niveau du complexe pyruvate déshydrogénase, c’est-à-dire avant le début réel du cycle de KREBS.
· Le second niveau de régulation se fait au niveau de la citrate synthétase. Elle est inhibée par l’ATP. En effet quand il y a assez d’ATP, il n’y a pas besoin d’en synthétiser.
· Le troisième niveau de régulation se fait au niveau de l’isocitrate déshydrogénase. Cette enzyme est catalysée par l’ADP.
· Le troisième niveau de régulation est le plus important et il se fait au niveau de l’alphacétoglutarate déshydrogénase. Cette enzyme est inhibée par le succinyl CoA et le NADH.
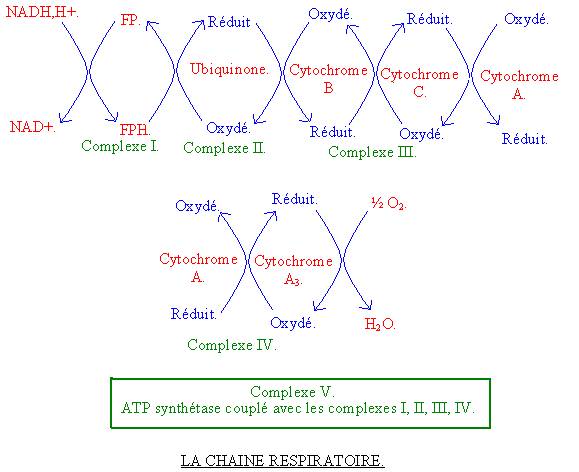
V – LA CHAINE
RESPIRATOIRE :
La chaîne respiratoire se passe dans la mitochondrie et plus particulièrement au niveau de la membrane interne de la mitochondrie. C’est le lieu le plus important de la resynthèse d’ATP. En effet les électrons arrachés aux substrats énergétiques qui ont été captés par les transporteurs d’électrons (NADH et FADH) vont être cédés à la chaîne respiratoire qui va les utiliser pour générer de l’énergie. Cette énergie permettra la resynthèse de nombreuses molécules d’ATP. Cette chaîne respiratoire qui utilise les électrons est composée de différents complexes, qui sont eux-même associé à un complexe enzymatique : L’ATP synthétase.
NADH et FADH sont produits au niveau du cycle de KREBS et de la bétaoxydation par la mitochondrie. Mais ils peuvent aussi venir du cytoplasme, car ils sont aussi produits par la glycolyse. Toutefois il y a un problème pour les NADH cytoplasmiques, car ils sont trop grands pou rentrer dans la mitochondrie. Afin de ne pas gaspiller les électrons, il existe un système, qui va permettre de transférer des électrons cytoplasmiques à la chaîne respiratoire. Ce système s’appelle la Navette-maltate / aspartate. Son principe repose sur le fait que les électrons cytoplasmiques sont cédés par le NADH cytoplasmique au maltate qui devient ainsi réduit dans le cytoplasme. Le maltate peut lui par contre entrer dans la mitochondrie grâce à un transporteur. Une fois dedans il cède ces électrons aux NAD mitochondriaux. Ce maltate oxydé va s’accumuler dans la mitochondrie. Or s’il s’y accumule, il n’y en aura plus dans le cytoplasme et la Navette ne fonctionnera plus. Il faut donc imaginer un système qui va faire sortir la maltate oxydé de la mitochondrie. C’est en fait un de ces composés très proche qui va sortir : L’aspartate. Les sorties vont être ainsi équilibrées aux entrées.
5-1 : LES
REACTIONS :
C’est une suite de réactions d’oxydo-réduction. Les électrons cédés par NADH et FADH à divers composés jusqu’à être captés par l’oxygène (oxygène inspiré). A certain niveau de cette chaîne de réaction d’oxydo-réduction suffisamment d’énergie est libérée pour permettre la synthèse d’une molécule d’ATP. Cette chaîne respiratoire est organisée sous forme de 5 complexes : 4 sont des complexes d’oxydo-réductions et 1 est le complexe où se fait la resynthèse d’ATP.
5-1-1 : Première
réaction :
![]() NADH
est oxydé sous forme de NAD et cède ces électrons au complexe 1, qui s’appelle
la flavoprotéine, qui devient donc réduite. Il y a suffisamment d’énergie
libérée à ce niveau pour permettre la resynthèse d’un ATP au niveau du complexe
5, qui s’appelle ATP synthétase : ADP + P ATP. C’est le premier ATP resynthétisé à partir de NADH.
NADH
est oxydé sous forme de NAD et cède ces électrons au complexe 1, qui s’appelle
la flavoprotéine, qui devient donc réduite. Il y a suffisamment d’énergie
libérée à ce niveau pour permettre la resynthèse d’un ATP au niveau du complexe
5, qui s’appelle ATP synthétase : ADP + P ATP. C’est le premier ATP resynthétisé à partir de NADH.
5-1-2 : Deuxième
réaction :
La flavoprotéine réduite est oxydée et cède ces électrons à l’ubiquinone, qui devient donc réduite. Elle correspond au complexe 2.
5-1-3 : Troisième
réaction :
L’ubiquinone réduite s’oxyde et cède ces électrons au complexe 3 et en particulier au cytochrome B qui devient réduit.
5-1-4 : Quatrième
réaction :
Ce cytochrome B cède ces électrons au cytochrome C, qui à son tour devient réduit. L’énergie est libérée au niveau de ce complexe 3 pour la resynthése d’une seconde molécule d’ATP par l’ATP synthétase.
5-1-5 : Cinquième
réaction :
Le cytochrome C réduit cède ces électrons au cytochrome A, qui devient à son tour réduit. Celui-ci cède alors ces électrons au cytochrome A3, qui est donc réduit. Au niveau de ce complexe 4, suffisamment d’énergie est libérée pour permettre la resynthèse d’ATP par le complexe 5. Le troisième ATP est resynthétisé.
5-1-6 : Sixième
réaction :
Le cytochrome A3 cède ces électrons à l’oxygène, en l’occurrence une demi-molécule d’oxygène. Au cours de cette réaction, il y est produit une molécule d’H2O, qu’on appelle eau métabolique.
Dans le cas de FADH, celui cède ces électrons au complexe 2 de la chaîne respiratoire. Par la suite c’est le même processus. Donc pour une molécule de FADH, il n’y aura resynthèse que de deux molécules d’ATP au lieu de trois pour NADH. Donc un NADH mitochondrial permet la resynthèse de trois ATP, et un FADH mitochondrial permet de resynthétiser deux ATP. Toutefois un NADH produit dans le cytoplasme ne permet de resynthétiser dans la chaîne respiratoire que deux ATP. Il a en effet du emprunter la Navette maltate / aspartate, qui va consommer une molécule d’ATP.
VI - BILAN
BIOENERGETIQUE DE LA DEGRADATION DUGLUCOSE :
On parle aussi d’oxydation du glucose.
|
Glycolyse. |
Substrat : Glucose sanguin. Produit : Acide pyruvique. |
+ 2NADH. |
0. |
+ 2 ATP. |
|
Transfert du pyruvate en acétyl CoA. |
Substrat : Acide pyruvique. Produit : Acétyl CoA. |
+ 2 NADH (2 * 1) |
0. |
0. |
|
Cycle de Krebs. |
Substrat: Acétyl CoA. Produit : Acétyl CoA. |
+ 6 NADH. |
+ 2 FADH. |
+ 2 ATP (GTP). (2 * 1) |
|
Chaîne respiratoire. |
Substrat : NADH et FADH : · 2 NADH (glycolyse): ·
2 NADH
(mitochondrie): ·
6 NADH (Cycle
de Krebs): ·
2 FADH
(Cycle de Krebs): |
0. 0. 0. 0. |
0. 0. 0. 0. |
+ 6 ATP (ou 4). + 6 ATP. + 18 ATP. + 4 ATP. |
Donc pour une molécule de glucose, on obtient 38 (ou 36) molécules d’ATP.
Le glucose et 6 O2 donne aussi 6 CO2 et 6 H2O.
![]() C6H12O6
+ 6O2 + 38 P + 38 ADP 6 CO2 + 6H2O + 38 ATP.
C6H12O6
+ 6O2 + 38 P + 38 ADP 6 CO2 + 6H2O + 38 ATP.
VII – LA BETA –
OXYDATION:
7-1 :
INTRODUCTION :
C’est la partie spécifique de la dégradation de l’acide gras. Elle se passe en partie dans le cytoplasme et en partie dans la mitochondrie. L’acide gras vient soit des triglycérides soit viennent du sang.
7-2 : LES
DIFFERENTES REACTIONS :
7-2-1 : Activation de
l’acide gras :
L’activation de l’acide gras se fait grâce à l’acide gras thiokinase. La réaction va donner de l’acyl CoA. L’acide gras est phosphoryle et le phosphate provient de l’hydrolyse de l’ATP.
![]() A.G + ATP + CoA Acyl CoA +
ADP.
A.G + ATP + CoA Acyl CoA +
ADP.
Il existe plusieurs types d’acide gras, mais tous suivent le même type de réaction.
7-2-2 : La pénétration
dans la mitochondrie de l’acyle Choa :
Ce composé ne peut entrer tel quel dans la mitochondrie. Un système de transfert va être nécessaire :
· L’acyle Choa est transformer en acyl carnitine. Cette réaction est catalysée par la CoA acyltransférase 1.
· L’acyl carnitine passe au travers des deux membranes de la mitochondrie grâce à un transporteur : La carnitine translocase.
· L’acyl carnitine est transformé dans la mitochondrie en acyl CoA. Cette réaction est catalysée par la CoA transférase 2. Il va y avoir accumulation de carnitine dans la mitochondrie qu’il faudra évacuer.
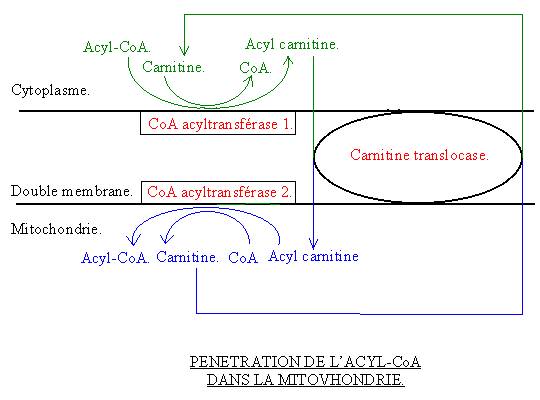
7-2-3 : La formation du
déhydroacyl-CoA :
On est alors dans la mitochondrie. La réaction est catalysée par l’acyl-CoA déhydrogénase. L’acyl CoA est oxydé et un FADH est réduit en FADH.
7-2-4 : Formation du
bétahydroxyacyl-CoA :
La réaction est catalysée par la déhydroxyacyl-CoA hydratase. Il va y avoir fixation d’une molécule d’H2O.
7-2-5 : Formation du
bétacétoacyl-CoA :
La réaction est catalysée par la bétahydroxyacyl-CoA déshydrogénase. Il y a oxydation du bétahydroxyacyl-CoA et une réduction du NAD.
7-2-6 : Formation de
l’acétyl-CoA :
Le bétacétoacyl-CoA se divise en deux molécules :
· Une molécule d’acétyl-CoA à deux carbones.
· Une molécule d’acyl-CoA ayant perdu deux carbones.
La réaction est catalysée par la bétacétothiolase. L’acétyl-CoA rejoint alors le cycle de Krebs et l’acyl-CoA fait autant de tour d’hélice nécessaire pour qu’il perde tous ces carbones. Il en perd deux par cycle.

VIII – BILAN
BIOENERGETIQUE :
Pour un acide gras, il y a un ATP consommé et par tour, il y a un FADH produit et un NADH produit. Le nombre de tours dépend de la taille de l’acide gras. Pour l’acide palmitique, il y a ainsi 129 ATP.
IX - EXERCICES:
1-1 :
PREMIERE EXERCICE :
Quel est l’équivalent énergétique d’un litre d’O2 dans le cas de l’oxydation du glucose ?
Quand le glucose est totalement dégradé, il y a libération d’énergie : 2839,9 KJ. Donc 6 O2 correspondent à 2839,9 KJ. Donc quand on consomme 6 molécules d’O2, il y a production de 2839,9 KJ. Une mole d’O2 fait 22,4 L.
Donc 6 * 22,4 L équivaut à 2839,9 KJ. Ainsi un litre d’O2 équivaut à 21 KJ.
Donc EEO2 (G) = 21 KJ.LO2-1.
1-2 :
DEUXIEME EXERCICE :
C’est le même exercice que le précédant mais cette fois dans le cas de l’oxydation de l’acide palmitique :
![]() C16H32O2
+ 23 O2 16CO2 + 16H2O
+ 100090 KJ.
C16H32O2
+ 23 O2 16CO2 + 16H2O
+ 100090 KJ.
23 moles d’O2 équivalent donc à 10090 KJ et une mole d’O2 correspond à 22,4 L.
Ainsi un litre d’O2 correspond à 19,6 KJ : EEO2 (AP) = 19,6 KJ.LO2-1.
1-3 : TROISIEME EXERCICE :
Quelle énergie est libérée par la dégradation totale du glucose si on consomme 30 L d’O2 ?
E = 30 * 21,1
E = 633 KJ.
1-4: QUATRIEME EXERCICE:
Quelle est l’énergie libérée par la dégradation totale de l’acide palmitique si on consomme 30 litres d’O2 ?
E = 30 * 19,6.
E = 588 KJ.
1-5: CINQUIEME EXERCICE:
On sait que l’énergie libérée à partir de l’oxydation du glucose est de 1266 KJ. Combien de litres d’O2 alors sont consommés ?
VO2 = D.E / EEO2.
VO2 = 1266 / 21,1 = 60 LO2.
VO2
= 60 * 22.4 = 1344 mol d’O2.
1-6: SIXIEME EXERCICE:
Quel est le volume d’O2 (VO2) nécessaire pour synthétiser 14 ATP par l’oxydation complète du glucose ?
![]() C6H12O6
+ 38 P + 38 ADP + 6 O2 38 ATP + 6 CO2.
C6H12O6
+ 38 P + 38 ADP + 6 O2 38 ATP + 6 CO2.
38 ATP correspondent à 6 oxygènes et une mole d’oxygène correspond à 22,4 L d’O2.
Donc 38 ATP correspondent à 6 * 22,4 L d’O2 et un ATP correspond 3,54 L d’O2. Donc 14 ATP correspondent à 49 L.
1-7 : SEPTIEME EXERCICE :
Combien de glucose est utilisé par une VO2 de 30 litres.
30 litres d’oxygène correspondent à 30 / 24 = 1,25. Donc un glucose correspond à 6 moles d’oxygène. Ainsi 30 litres d’O2 correspondent à 0,22 mol de glucose.
Or une mole de glucose correspond à 180 grammes ; Donc M(Glu) = 39,6 g.
Donc quand on consomme 30 litres d’oxygène, on oxyde 39,6 g de glucose.
1-8 : HUITIEME EXERCICE :
Combien de temps en heure, en minute et en seconde mettons – nous pour oxyder tout le stock de glucose quand on oxyde le glucose. Avec une VO2 en 30 minutes de 30 litres et un stock de glucose de 39,6 g ?
On sait qu’avec 30 litres d’O2, on consomme 39,6 g de glucose.
Donc un litre d’O2 par minute correspond à 2,6g (Glu).min-1.
On en a donc pour 1h 15 min et 42 s.