3.3 Mémoire déclarative ou explicite


![]()
![]()
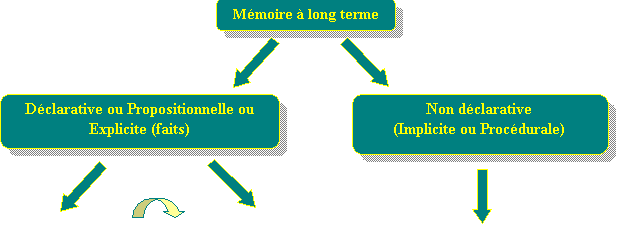
Un modèle d'organisation de la mémoire à long terme
Les tests de mémoire explicite mettent en jeu des épreuves de rappel (comme dans l'expérience de Godden et Baddeley, 1975) ou des épreuves de reconnaissance (par exemple, distinguer un événement ou une information au milieu d'autres événements ou informations). "Elle fait appel à des processus conscients, intentionnels et verbalisables". "Elle est donc la mémoire des souvenirs conscients et durables" aisément accessibles, que le sujet peut rapporter, restituer. Elle comprend les connaissances et les savoirs que l'étudiant ou le professionnel a su se construire. L'accès à la mémoire explicite est rapide.
La mémoire déclarative ou explicite se subdivise ensuite selon Tulving (1985) en deux mémoires ou systèmes de mémoire, la mémoire sémantique et la mémoire épisodique. Selon les uns ou les autres, la mémoire sémantique peut (Tulving) ou non fonctionner indépendamment de la mémoire épisodique. Cet auteur attribue en outre des niveaux de conscience à chacun des différents (3) systèmes de mémoire. Ainsi, à partir du terme grec noesis (savoir) il associe la mémoire épisodique à la conscience autonoétique (conscience de soi), la mémoire sémantique à la conscience noétique (conscience de savoir) et la mémoire procédurale à la conscience anoétique (inconscience).
a) Episodique
Parfois appelée autobiographique dans la mesure où elle fait référence à l'individu soit en tant qu'acteur, soit en tant qu'observateur des événements mémorisés, la mémoire épisodique est une mémoire des événements biographiques inscrits dans un contexte donné. C'est une mémoire égocentrée qui offre une perspective singulière et subjective sur des événements vécus par un individu et non rapportés par autrui. Elle correspond à la mémoire de ses expériences personnelles (les souvenirs de l'enfance ou du dernier réveillon par exemple, ou bien le lieu de vacances de l'été dernier), avec des éléments relatifs à ce contexte (le moment, le lieu, les personnes présentes, autant d'éléments qui pourront servir d'indices de récupération du souvenir). Elle est en perpétuelle évolution car de nouveaux événements viennent s'ajouter à chaque moment de notre vie.
b) Sémantique
La mémoire sémantique permet à l'individu de stocker sa connaissance du monde. Elle nous permet de retrouver des informations indépendamment du contexte dans lequel nous en avons fait l'acquisition (nous ne savons pas nécessairement quand ni à quelle occasion nous avons appris ce que nous savons). Je peux ainsi savoir que Victor Hugo a écrit Les Misérables sans jamais avoir rencontré Victor Hugo ou lu cet ouvrage. Le contenu de cette mémoire nous est accessible rapidement, facilement, sans effort. Mémoire sémantique signifie bien sur mémoire du sens des mots, elle comprend essentiellement l'information nécessaire à l'utilisation du langage (l'étude des troubles du langage permet l'étude de la mémoire sémantique). Elle permet à l'individu de disposer également d'un certain nombre de connaissances générales (la terre est ronde, Paris est la capitale de la France) accumulées tout au long de son existence et structurées. Elle peut être définie comme ensemble de connaissances objectives, de faits ou d'événements qui se sont déroulés indépendamment de moi. Elle rend possible les représentations cognitives et le classement des objets, des situations, des faits, des événements. Contrairement à la mémoire épisodique, la mémoire sémantique demeure relativement stable, solide. Elle est moins affectée par différents traumatismes que ne l'est la mémoire épisodique.
Mémoire épisodique et mémoire sémantique ne sont pas des entités isolées mais interagissent constamment l'une avec l'autre. L’opposition entre ces deux types de mémoire est sans doute réductrice. Il existe vraisemblablement un lien étroit entre la mémoire épisodique et la mémoire sémantique. En effet, la mémoire épisodique permet non seulement de se souvenir d’expériences spécifiques, mais encore de développer la mémoire sémantique qui permet l’accès à la connaissance du monde. De son côté, la mémoire sémantique peut être considérée comme le produit des diverses expériences stockées dans la mémoire épisodique. Elle permet de souligner en les décontextualisant les traits communs aux divers épisodes. Une transition progressive s'effectue donc de la mémoire épisodique à la mémoire sémantique, en permettant favorisant un processus de généralisation.
II
– Les structures et les circuits de la
mémoire.
1) Plasticité neuronale et
potentialisation à long terme (LPT)
Nous avons vu que la mémoire était le fruit d'un processus actif, dynamique. Pour la plupart des auteurs, la mémoire est à la fois focale, locale, et en même temps largement distribuée. Ceci signifie que des aires distinctes traitent et stockent des aspects différents d'une même représentation. De petits ensembles neuronaux traiteraient et stockeraient des composantes particulières de l'information mais la représentation d'un événement dans sa globalité nécessiterait la présence de nombreuses aires (plusieurs systèmes mnésiques).
La notion de réseau neuronal est tout à fait centrale dans cette conception, correspondant à un modèle de représentation connexionniste. En effet, aucun neurone isolé ne contient en lui-même l'information nécessaire à la restitution d'un souvenir. L'existence de la trace mnésique ne peut être mise en évidence que lorsque qu'un réseau de plusieurs neurones interconnectés est activé. Selon Changeux (1983), la représentation mentale est l'état physique créé par l'entrée en activité (électrique ou chimique), corrélée ou transitoire, d'une large population de neurones distribués. La mémorisation d'un souvenir ou d'un événement, résulte de la modification, de la réorganisation permanente des différentes connexions neuronales spécifiques mises en jeu et activées lors du traitement d'une information permettant l'évocation le rappel ou la restitution de ce souvenir. Un circuit neuronal n'est pas dédié à l'évocation d'un souvenir en particulier, différents souvenirs peuvent être encodés à l'intérieur d'un même réseau de neurones par différentes configurations ou connexions synaptiques. Mais un souvenir peut aussi de la même façon faire appel à la mise en jeu simultanée de plusieurs assemblées de neurones distribuées, réparties dans différentes aires du cerveau.
Cette activité simultanée de neurones au sein de différents réseaux confère au cerveau des possibilités extraordinaires se caractérisant notamment par sa capacité à s'adapter, à modifier, à remodeler ses propres réseaux. Ceci témoigne en définitive d'une grande plasticité synaptique ou neuronale, mécanisme de base du fonctionnement de la mémoire. Cette plasticité cérébrale se manifeste principalement par un mécanisme appelé potentialisation à long terme (LTP), découvert dans l'hippocampe en 1973, mais dont la présence a par la suite été démontrée dans de nombreuses régions du cortex. Pourquoi parle-t-on de potentialisation à long terme ? La potentialisation (ou l'activation, l'excitation neuronale) survient à la suite d'une expérience sensorielle qui va produire une trace mnésique par la modification induite dans le circuit neuronal. Certaines synapses activées, sollicitées, vont être ainsi renforcées, ce processus donnant naissance à des modifications importantes et durables (création et stabilisation de nouveaux réseaux, permettant l'inscription en mémoire de cette expérience).
|
|
|
|
|
|
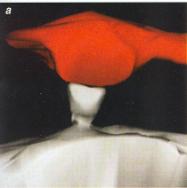
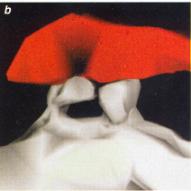
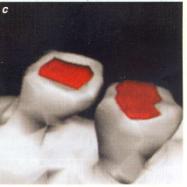
Ces images de synapses ont été obtenues par la reconstruction
tridimensionnelle. Lorsque la synapse n'est pas activée, la connexion entre
le neurone présynaptique, où arrive l'influx nerveux, et le neurone
postsynaptique n'est constituée que d'une épine unique (a). |
Quand la synapse est activée, une deuxième épine
"pousse" (b). De surcroît, les synapses doubles qui se forment
possèdent toutes deux les récepteurs nécessaires à la transmission du signal
(zones rouges sur la photographie). (c) : elles sont vraisemblablement
fonctionnelles. |
||
|
(Pour la science, 2001) |
|||
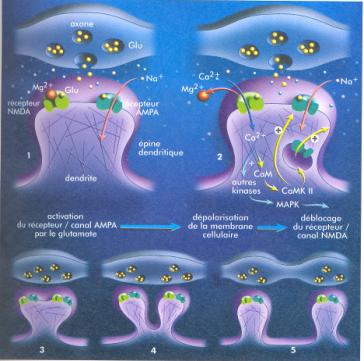
Ainsi, lors de la survenue d'une nouvelle expérience sensorielle dans les mêmes conditions déjà connues, les mêmes cellules seront activées. Les neurones réagissent davantage à toute excitation ou activation ultérieure par ces mêmes voies nerveuses. Cette potentialisation à long terme se caractérise par sa persistance qui peut durer plusieurs mois. Il existe un phénomène inverse de la LTP, la dépression à long terme (LTD), qui entraîne un silence de la synapse. Cette dépression joue un rôle important de correction au niveau de la mémoire procédurale (les réseaux neuronaux impliqués dans des mouvements erronés sont inhibés par la mise sous silence de leurs connexions synaptiques). La mémorisation se fonde donc sur des mécanismes moléculaires mettant en jeu des neuromodulateurs, des neuromédiateurs. Le glutamate, neurotransmetteur libéré dans la fente synaptique séparant le neurone pré synaptique qui a reçu l'excitation sensorielle du neurone post synaptique, va se fixer sur plusieurs types de récepteurs différents dont deux sont particulièrement importants pour la LTP : les récepteurs AMPA et NMDA. Le récepteur AMPA est couplé à un canal ionique qui provoque l'entrée de sodium dans le neurone post-synaptique lorsque le glutamate s'y fixe. Cette entrée de sodium amène la dépolarisation locale du dendrite et la transmission de l'influx nerveux dans le neurone suivant, si cette dépolarisation atteint le seuil de déclenchement du potentiel d'action.
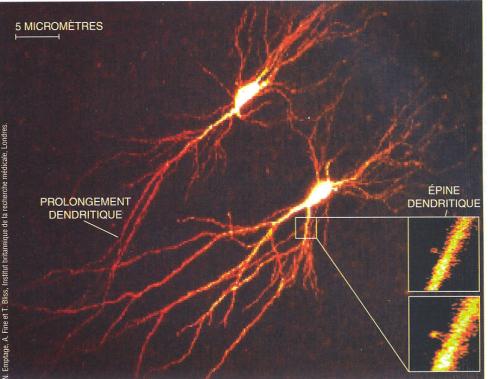
|
|
||
|
L'activation de deux neurones a été visualisée après injection d'un marqueur fluorescent des ions. L'image d'une portion de dendrite avant activation synaptique (cartouche en haut à droite) et pendant l'activation (en bas à droite) montre l'élévation transitoire de la concentration en ions calcium (en jaune) dans une épine dendritique et dans un prolongement. Les ions calcium activent la synthèse de diverses protéines utilisées pour le renforcement des protéines. (Pour la science, 2001) Quand le récepteur NMDA, également un récepteur couplé à un canal
ionique, est activé, des ions calcium pénètrent dans le neurone post
synaptique. Cette
concentration accrue de calcium dans le dendrite active toute une série de
protéines, des kinases qui vont déclencher l'expression de divers gènes
codant des protéines essentielles à la modification de la synapse, à
l'augmentation prolongée de son efficacité et à la construction de nouvelles
synapses.
|
Nous terminerons sur ce point en rappelant que les aspects neurophysiologiques de la mémoire que nous venons d'évoquer sont nécessaires mais pas suffisants pour en expliquer le fonctionnement, notamment dans une perspective psychologique ou cognitiviste. La connaissance du système nerveux ne suffit pas à répondre aux nombreuses questions que posent à la fois la complexité et l'organisation de la mémoire.
2) Le rôle de l'hippocampe
a) Un rôle essentiel dans la construction de la mémoire
On sait depuis 1953 que les différents types de mémoire font appel à des circuits neurobiologiques distincts. L'histoire originale et tout à fait incontournable lorsque l'on aborde cet aspect a été rapportée par William Scoville (opérant aux Etats-Unis) et Brenda Milner en 1953. Il s'agit du patient H.M., opéré à 27 ans afin de le soulager de graves crises d'épilepsie. L'opération a constitué à enlever de façon bilatérale certaines parties du cerveau dont l'hippocampe, l'amygdale (ce nom provient d'un emprunt ancien au latin –XIIIes.-, du grec amugdalê qui signifie amande), et une partie du lobe temporal. Ceci a permis au patient de récupérer de l'épilepsie, mais a provoqué l'apparition d'amnésies antérogrades profondes. Ainsi, bien qu'il ait conservé un empan normal, H.M. a commencé d'oublier tous les événements de sa vie quotidienne, perdant tout capacité de former de nouveaux souvenirs. Si sa mémoire explicite était très touchée, en revanche sa mémoire implicite semblait préservée. Par exemple, à chaque présentation du test de la tour de Hanoï, le patient ne se rappelait pas l'avoir déjà vu (ce qui confirme que l'intégralité du lobe temporal médian est absolument indispensable au bon fonctionnement de la mémoire déclarative) mais améliorait ses performances de jour en jour (procédurale – le striatum est très actif dans les apprentissages procéduraux).
|
|
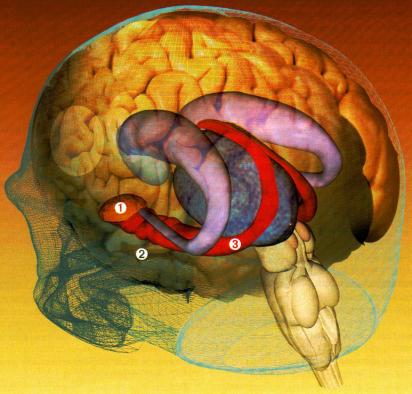
L'hippocampe, impliqué dans l'acquisition et le stockage des souvenirs épisodiques, est situé dans le lobe temporal médian, près de l'amygdale. |
Le rôle de l'hippocampe et des formations para hippocampiques (amygdale, formation hippocampe, cortex parahippocampique (rhinal entorhinal) dans la formation et la préservation de la mémoire a ainsi été mis en évidence, l'ablation de ces structures provoquant certains types d'amnésies.
L'hippocampe est constitué de deux régions distinctes, le gyrus denté et la Corne d'Ammon (en raison de la ressemblance de cette structure avec la forme des cornes de bélier d'Amon, Dieu de Thèbes) qui comprend différentes aires constituées de neurones pyramidaux, les aires CA1, CA2 et CA3 (l'acronyme CA signifie Corne d'Ammon). Des singes présentant des lésions spécifiques de la région CA1 montrent des troubles des mémoires antérograde et rétrograde (des connexions antérogrades convergent vers l'hippocampe en même temps que des connexions rétrogrades en partent vers les cortex primaires pour y stocker et consolider, via les synapses corticales, les associations facilitées par l'hippocampe). Les informations uni ou multimodales issues des différentes aires sensorielles (visuelle, auditive, etc.) vont converger et parvenir à l'hippocampe en passant par différents cortex (rhinal, entorhinal – le cortex rhinal avait été également retiré chez le malade H.M.), créant différentes connexions qui vont former ce qu'on appelle la boucle tri synaptique de l'hippocampe qui sert probablement à encoder la mémoire spatiale.
Il semble qu'existe, en fonction des différentes mémoires (tâche de reconnaissance visuelle, auditive, tactile, etc.) différents circuits incluant et reliant par des connexions réciproques les lobes temporaux (l'hippocampe est situé sur la face interne), des structures sous corticales (comme l'amygdale), le septum, l'hypothalamus (ces différentes régions reliées entre elles font partie, avec le bulbe olfactif,les corps mamillaires, etc. de ce qu'on appelle le système limbique) et d'autres régions extra limbiques comme les aires associatives et le cortex frontal. Grâce à ces connections neuronales, le cortex et le système limbique entretiennent un échange très riche d'informations, indispensable à la mémorisation, à l'apprentissage et aux comportements affectifs de l'individu.
|
|
|
|
Le système limbique : A- corps calleux, B- voie olfactive, C- corps
mamillaires, D- fornix, E- noyaux antérieurs du thalamus, F- amygdale,
G- hippocampe,
H- gyrus parahippocampique, I- circonvolution cingulaire, J- noyaux
hypothalamiques (Source schéma légendé : Le cerveau à tous les niveaux) |
|

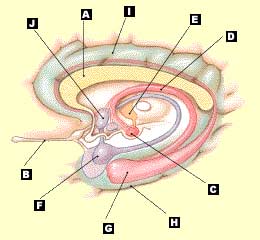
Le système limbique participe à la construction de la mémoire car il intervient par exemple dans les aspects émotionnels, grâce notamment à l'amygdale. L’amygdale joue un rôle essentiel dans la genèse de l’expression des émotions. Meunier (2001) et Berthoz (2003) rappellent que "c'est à Heinrich Klüver et à Paul Bucy (Université de Chicago, 1938), que l'on doit d'avoir observé chez le singe les effets d'une lobectomie bitemporale (plus précisément la région antérieure de la partie médiane) incluant les deux amygdales. Ceci induit un syndrome (dit depuis de Klüver et Bucy) caractérisé par un comportement hypersexuel et une hyperoralité. L’animal mange sans discrimination même des aliments qu’il aurait rejetés, et surtout il manifeste une placidité remarquable devant les événements les plus effrayants et aucune réaction de peur." D'autres expériences ont montré depuis l'incapacité pour des singes présentant une lésion bilatérale de l’amygdale d'associer la notion de récompense ou de punition dans différentes tâches. De façon générale, conclut Berthoz, "il est maintenant clair que l’activité de l’amygdale est, chez l’homme, liée à l’activité mentale d’évocation de souvenirs lointains à caractère émotionnel et donc influence la mémoire émotionnelle."
Le circuit de Papez, ou circuit hippocampo-mamillo-thalamique représente un des réseaux reliant les différentes structures limbiques entre elles. Une lésion de ce circuit peut être à l'origine d'un trouble ou d'un syndrome particulier. Il est en ainsi du syndrome de Korsakoff (cf. le TD pour ce syndrome alcoolo-carentiel pour reprendre l'expression d'Eustache et Faure, i.e. alcolisme prolongé et carence alimentaire notamment en vitamine B1) caractérisé entre autres par une amnésie antérograde, c'est-à-dire ne permettant plus la formation de nouveaux souvenirs. L'information à mémoriser parcourt les différentes structures du circuit de Papez, de l'hippocampe aux corps mamillaires de l'hypothalamus, au noyau antérieur du thalamus, etc. avant de revenir à l'hippocampe. Ce cheminement de l'information entraîne un remodelage physique des circuits nerveux, favorise la consolidation de cette information et permet de construire progressivement son indépendance vis à vis de l'hippocampe (la lésion de cet organe n'empêchant pas le stockage et le rappel des souvenirs encodés avant la lésion). L'hypothèse est que les souvenirs se trouvent peu à peu stockés dans les zones spécifiques du système neuronal correspondant aux régions sollicitées au moment de la perception, du traitement et de l'encodage de ces souvenirs (auditif, visuel, etc.). La réactivation de l'ensemble des éléments du réseau permet la restitution intégrale du souvenir. "Une des fonctions importantes de l'hippocampe serait donc de détecter et de mémoriser des combinaisons simultanées d'indices sensoriels multiples. L'hippocampe serait critique pour l'apprentissage de la conjonction d'indices perceptifs simultanés, procédant à la recomposition des épisodes multisensoriels à partir d'éléments incomplets" (Berthoz, 2003).
Certains chercheurs ont émis l'hypothèse (cohérente avec ce que nous venons d'évoquer) d'un rôle certes essentiel mais transitoire de l'hippocampe dans le processus de mémorisation. Cet organe jouerait un rôle temporaire dans un mécanisme de consolidation rapide au cours duquel il mettrait en relation les informations initialement enregistrées dans le cortex. Ce mécanisme mettrait en jeu des connexions réciproques et permettrait donc un dialogue entre l'hippocampe et les aires néocorticales. En effet, quelques temps après l'ablation de cette structure, (Zola-Morgan et Squire, en 1990 à San Diego), la lésion n'a plus d'effet sur la remémoration de ce qui a été appris. L'hippocampe participerait initialement au stockage et au rétablissement de la mémoire déclarative, puis peu à peu se mettrait en place un mécanisme de consolidation et de réorganisation de façon à établir une mémoire permanente. (Pour la science Potier, Billard et Dutar).
|
|
|
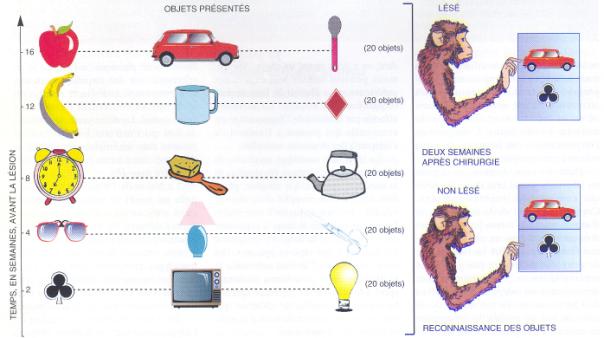
Un très grand nombre d'objets ont été
présentés aux singes sur une durée de 2 à 16 semaines. Certains singes
subissent ensuite une ablation de l'hippocampe. Après cette ablation, les
singes lésés se rappellent mieux les souvenirs les plus anciens alors que les
singes non lésés se remémorent mieux les objets vus le plus récemment (Source
Potier et coll, Pour la Science, 2001). |