A : Relation tension active-longueur :
Cette relation est en fait une parabole, c’est-à-dire que la tension passe par un maximum pour une longueur du muscle qui est très proche de sa longueur au repos et qui diminue progressivement quand le muscle se raccourcie ou augmente.
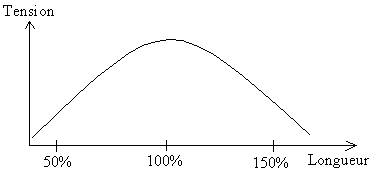
Les résultats montrent que la force dépend de la longueur des sarcomères : La tension est maximale quand la longueur est proche de 2 ou 2,4, car c’est à ce niveau que le chevauchement des filaments fins et épais est optimal (maximum de liaisons actine-myosine). La force dépend donc du nombre de liaisons entre l’actine et la myosine. Si on stimule le muscle quand il est étiré, il y a peu de liaison et donc peu de force. (150% de sa longueur de repos) ; Et inversement.
B : Relation tension globale-longueur :
Au niveau du muscle entier, il faut tenir compte des éléments d’élasticité : Si on n'intègre pas les tendons, les résultats vont être différents.
La tension globale est la résultante des tensions actives et de la tension passive. Elle se superpose à celle active quand le muscle n’est pas en allongement (longueur de repos au plus faible). Par contre la tension globale augmente avec la longueur du muscle : Ca atteint un maximum quand ça atteint une longueur légèrement supérieure à la longueur de repos. Cela s’explique par l’intervention passive qui diffère de 0 quand la longueur est supérieure à 100%.
On considère que la marge de manœuvre du muscle est entre 80 et 130% de sa longueur initiale. La force maximale d’un muscle sera obtenue pour une longueur légèrement supérieure à la longueur de repos. Donc si le muscle est préalablement étiré, la force de contraction sera plus grande. Ceci explique par la répartition de l’énergie élastique lors de l’étirement. Il y a ainsi transmission d’énergie au muscle pendant la contraction. La force est alors plus grande.

Ce processus de stockage et de restitution est impliquée dans de nombreux mouvements, comme la marche et la course. Ca permet de diminuer le coût énergétique.
C : Relation tension globale-force :
Le tissu conjonctif influence la force passive. Sa quantité va être selon la proportion de fibres dans un muscle. Un muscle qui a beaucoup de filaments et donc beaucoup de collagène. Un muscle lent est plus raide qu’un muscle rapide. : La compliance est plus faible.
Le pourcentage de fibres joue un rôle sur la diminution de la force maximale : Un muscle lent aura un plateau assez large, mais s’il contient beaucoup de fibres rapides, il n’y a pas de plateau (plage d’action plus faible). Ca dépend aussi de la disposition des fibres dans le m du nombre muscle.
5-3-4 : Relation force-vitesse :
Elle montre que la force développée par un muscle est différente selon sa vitesse. On impose une vitesse au muscle et on regarde la force. Cela montre que la vitesse maximale de raccourcissement dépend de la charge soulevée.
La vitesse est maximale quand la charge est nulle (ou la force est nulle) et inversement : La force produite est maximale quand la vitesse de raccourcissement est nulle (contraction isométrique). Entre ces deux extrêmes, la vitesse diminue régulièrement quand la force augmente. La force maximale développée sera donc plus faible quand le muscle sera en contraction concentrique pure par rapport à la contraction isométrique.

Quand le muscle s’allonge, on observe l’inverse : Plus la vitesse d’allongement est grande et plus la force développée sera aussi grande. Aussi l’augmentation de la vitesse d’allongement et la force sera grande, jusqu’à un plateau. Il arrive à un moment donné où si la vitesse augmente, la force diminue.

Pour une même vitesse, la force développée en excentrique (raccourcissement) est plus grande qu’en excentrique. Cela s’explique par la durée de la liaison actine-myosine : Plus la vitesse de raccourcissement augmente, moins de liaisons se formeront et on diminue la durée de ces liaisons. Ca implique la diminution de la force développée au niveau de chaque liaison et au niveau total du muscle.
En excentrique c’est différent car les têtes de myosine s’inversent dans le sens inverse (Donc de plus en plus long).
Cette relation force-vitesse connaît des variations selon les muscles :
· La valeur de la force maximale : La force maximale dépend du diamètre du muscle et de la propulsion fibres lentes et rapides.
· La valeur de la vitesse maximale : Elle dépend de la longueur du muscle et dépend aussi de la proportion des fibres lentes et rapides : Plus il y a des fibres rapides et plus la vitesse sera grande.
La relation dépend aussi de la température. Le refroidissement d’un muscle diminue sa vitesse de raccourcissement et diminue aussi (mais moins) la force maximale produite.
La puissance (F * Vitesse) joue aussi. La relation qu’on vient de voir montre donc que quand la force est maximale, la puissance est nulle. C’est dans une zone située entre 30 et 50% de la vitesse maximale et de la force maximale que la puissance sera maximale.
La vitesse de contraction qui fournit cette puissance maximale est appelée la vitesse optimale.
5-4 : LA FORCE MUSCULAIRE
MAXIMALE :
La force qui peut être développée par un muscle est liée au nombre de liaisons crées. Donc tous les facteurs qui influencent le nombre de liaisons, influencent aussi la force.
5-4-1 : Mécanisme de graduation de la force
musculaire.
Comment peut-on augmenter la force dans un mouvement ?
L’augmentation de la force musculaire fait intervenir principalement deux mécanismes : L’augmentation de la fréquence de décharge des unités motrices et l’augmentation du nombre des unités motrices. On enregistre cela grâce à l’EMG.
A : Augmentation du nombre d’unités motrices
actives :
C’est ce qu’on appelle le recrutement spatial. Un muscle comprend un grand nombre d’unités motrices : Plus il y aura d’unités motrices activées et plus la force musculaire développée sera grande. Toutes les unités motrices ne sont pas activées en même temps. Il y a un recrutement des unités motrices, qui augmente avec la force à produire. Si la force est faible, il y a activation de quelques unités motrices. Plus la force augmente et plus il y a activation d’unités motrices. C’est le recrutement spatial. Il n’est pas aléatoire. Il se fait selon un ordre précis :
· Les premières unités motrices activées sont les unités motrices lentes.
· Ensuite les unités motrices rapides : 2a puis 2b.
Aussi quand toutes les unités motrices du muscle sont activées, c’est à ce moment que le muscle développe sa force maximale. Ce recrutement s’explique par la taille du motoneurone. Les fibres lentes passent en premier car la taille du motoneurone est petite (diamètre petit) : Il est du coup plus excitable (seuil de dépolarisation plus bas) : Elles sont excitées en premier. Les fibres situées dans les motoneurones plus gros sont moins excitables donc sont recrutées plus tard.
Il existe un seuil de recrutement ou seuil d’activation qui correspond à la valeur de la force pour laquelle la valeur de l’unité motrice est mise en jeu dans le sens où il y a un lien direct entre l’amplitude de la secousse de l’unité motrice et son seuil de recrutement.
Les unités motrices de type 2 développent plus de force que les fibres de type 1 car :
· Les fibres rapides ont un diamètre plus grand et ont donc plus de myofibrille et plus de protéines contractiles. Elles peuvent donc faire plus de liaisons.
· Les fibres rapides contiennent plus de fibres.
Ce recrutement spatial des unités motrices est différent selon la vitesse de contraction du muscle. Lors d’une augmentation rapide de la force, il y a un recrutement rapide des unités motrices : Plus la vitesse est grande et plus le seuil de recrutement sera faible. Aussi il existe une alternance des activités des unités motrices. Si l’unité motrice vient d’atteindre son seuil, elle ne sera pas active continuellement. Ca ralentit le processus de la fatigue musculaire.
B : Augmentation de la fréquence de décharge de
l’unité motrice active :
C’est le recrutement temporel. Une fibre peut être stimulée avec une seule stimulation, mais un muscle peut être activé par plusieurs. Plus la fréquence de décharge des motoneurones sera grande et plus les potentiels d’action reçus par les fibres auront une fréquence importante. Et quand plusieurs stimulations se suivent rapidement, ils activent des fibres musculaires qui s’additionnent.
La fréquence de décharge n’est pas constante et varie selon le niveau de la force : Plus elle est grande et moins la fréquence sera faible. Donc les unités motrices proches de leur seuil d’activation déchargent de façon irrégulière. La fréquence de décharge optimale dépend de l’unité motrice. La fréquence maximale pour les fibres lentes est comprise entre 7 et 12Hz. La fréquence maximale pour les fibres rapides est de 100 à 150Hz.
Les déchargements des motoneurones ne sont pas forcément synchrones. Si la force est inférieure à 50% de la force maximale, les motoneurones des différentes unités motrices du muscle ne sont pas synchrones. Par contre plus la force sera élevée et plus les activités des unités motrices auront tendance à se synchroniser. Cette fréquence de décharge (= Recrutement temporel) joue un rôle sur la fréquence du muscle. Le recrutement temporel ne joue qu’un rôle secondaire par rapport au recrutement spatial.
C : Modification de l’activité des unités
motrice :
Lors de contraction prolongée, on observe que les recrutements sont séparer. On distingue les maintiens d’une force maximale et sous maximale :
· Maintien d’une force maximale :
- Modification de la fréquence de décharge : Certaines sont de plus en plus faible, d’autres vont augmenter leur fréquence de décharge et d’autres ne vont pas les modifier.
- Augmentation progressive du nombre d’unités motrices activées (modification du recrutement spatial) : Dès la première seconde de maintien.
· Maintien d’une force sous maximale : Diminution du nombre d’unités motrices actives et une diminution de leur fréquence de décharge. Et l’évolution sera différente selon les unités motrices :
- Les unités motrices dont la fréquence de décharge est faible garderont une activité tonique pendant le maintien mais la fréquence sera plus faible (de l’ordre de deux fois).
- Les unités motrices, dont la fréquence de décharge est grande, s’arrêtent et repartent par la suite.
Par conséquent quand on maintien une force, il y a fatigue des unités motrices. Les unités motrices fatiguées développent des forces de plus en plus faibles et des secousses dont le temps de relaxation sera de plus en plus long.
Quand on maintien une force sous maximale, il y a donc le recrutement de nouvelles unités motrices, pour compenser la perte de force. Quand on maintien une force maximale, toutes les unités motrices sont déjà actives. La diminution de la fréquence de décharge permet de limiter la diminution de la force, car il y a allongement du temps de relaxation et ceci fait que l’addition des secousses se fait pour des fréquences plus faibles, car la durée de la relaxation est augmentée. Par contre l’arrêt de l’activité des unités motrices les plus fatigables provoquent la diminution de la force. Il n’y a don pas de compensation possible.
5-4-2 : Influence du type de muscle :
A : Section et proportion des différents types
de fibres :
Pour un muscle donné, la force maximale développée est proportionnelle à son diamètre ( environ 22 N.cm2). Cette force maximale exprimée par section transversale de muscle est la force maximale spécifique. Pour une section transversale de muscle, la force maximale spécifique est d’autant plus élevée que la proportion de fibres 2 rapides va être importante.
B : L’arrangement architectural des
fibres :
Il existe deux grands types de muscles :
· Muscles fusiformes dans lesquels, les fibres sont parallèles.
· Muscles penniformes (muscles pennés) dans lesquels il y a une insertion latérale des fibres.
Les muscles fusiformes sont caractérisés par le fait qu’ils ont les fibres et les tendons dans la même direction et les fibres sont en parallèle sur toute la longueur du muscle. Ils sont rares et se trouvent au niveau des extrémités. Ils sont longs et fins, c’est-à-dire constitués de fibres longues mais peu nombreuses.
Les muscles penniformes sont caractérisés par des fibres qui s’implantent en obliques sur le tendon. C’est la majorité des muscles. Il y a plusieurs types de configurations :
· L’arrangement unipenné où les fibres s’implantent de façon oblique sur le tendon dans un sens.
· L’arrangement bipenne où les fibres s’implantent dans deux sens.
· L’arrangement multipenné.
Les fibres sont toujours en diagonale par rapport au tendon.

Pour un même volume de muscle, le nombre de fibres est plus important au niveau des muscles penniformes mais la longueur est plus courte. L’arrangement a une influence sur l’application de la force et l’amplitude de l’allongement de raccourcissement : En effet plus l’angle entre les fibres et le tendon est petit et plus le nombre de fibres pas unité de volume sera grande. Alors le nombre de liaisons sera plus grand et il y aura plus de force. La force est produite dans la direction des fibres et si l’angle est grand, seul une petite partie de la force sera donnée au tendon. Il existe un angle optimal de 30° pour que la force soit maximale. Si l’angle est supérieur, la force diminue.
Plus l’angle entre le muscle et les tendons est grand et plus la longueur est faible et plus l’amplitude de contraction est réduite. Donc les muscles impliqués dans des mouvements de grande amplitude développent peu de force et ce sont des fibres fusiformes, mais les muscles impliqués dans des mouvements de faible amplitude développent peu de force et ce sont des fibres penniformes.
5-4-3 : Influence des conditions de
stimulation :
Elle dépend de la longueur initiale du muscle. La force est maximale à une longueur qui est sa longueur de repos quand il est dans l’organisme (déséquilibre : Légèrement étiré).
Le type de contraction du muscle : Pour une même activation nerveuse, la force développée sera plus importante lors d’une contraction excentrique (en allongement) cumulée à une concentrique.
La vitesse de contraction : Elle agit sur la force que développe un muscle. Elle est d’autant plus faible que la vitesse de raccourcissement est grande. Donc la force maximale est développée en isométrie, et en allongement la force développée augmente avec la vitesse.
5-4-4 : Influence des facteurs biologiques,
physiologiques et psychologiques :
A : Facteurs biologiques :
· Effet du sexe, à savoir que les muscles des femmes développent moins de force que les hommes : A taille égale la force de la femme représente environ 75 à 80% de la force de l’homme. Au poids maigre les différences s’annulent. La plus faible force maximale spécifique des femmes (15%) s’explique par une présence de masse grasse plus grande.
· Effet de la latéralité (droitier ou gaucher) : On développe une force 5 à 6% supérieur par rapport au bras faible. De même on développe une force 8 à 9% supérieure par rapport à la jambe faible.
· Effet de l’âge : La force maximale augmente jusqu’à 20 ans, puis reste stable jusqu’à 30 ans puis diminue. La diminution devient manifeste à 50 ans. La force maximale spécifique est maintenue jusqu’à 80 ans. La diminution de la force est donc seulement due à une atrophie musculaire qui s’explique par :
- Une diminution du nombre des fibres, liées à la dégénérescence du motoneurone (ceux de grandes tailles sont les plus touchés, c’est ceux qui innervent les fibres rapides). C’est donc surtout une diminution des fibres rapides.
- Une diminution du diamètre des fibres 2. Le diamètre des fibres 1 reste constant.
Les effets de l’âge sur le muscle squelettique sont très liés à une diminution de l’activité physique : Les fibres 2 non sollicitées disparaissent et les fibres 1 sont sollicitées pour la posture. A 80 ans il y a une dégradation accentuée par la dégradation des propriétés des fibres contractiles.
B : Facteurs physiologiques :
Cette force musculaire varie selon la position de l’activation et de la position des articulations adjacentes.
Le degré de solidification du muscle : Plus il est solide et plus la force est grande. L’augmentation de la force par l’entraînement est liée au principe de surface, c’est-à-dire que le muscle s’adaptera que si la force qui lui est donnée est supérieure à la force traditionnelle. Il faut 3 à 4 entraînements pour noter une augmentation de force sur la première semaine. Cette augmentation implique deux facteurs :
· Un facteur nerveux, qui est responsable de l’augmentation initiale (au début de l’entraînement : 4 premières semaines). Ils correspondent à l’augmentation de l’activation nerveuse des fibres musculaires :
- Elle comprend une modification du recrutement des unités motrices. L’entraînement modifie la synchronisation des unités motrices.
- La diminution de l’activation des propriocepteurs musculaires et plus précisément des organes tendineux de Golgi (au niveau de la jonction tendon – muscle et envoie des informations au motoneurone quand l’étirement est trop grand.
- L’amélioration de la coordination musculaire dans le sens où il y a une diminution de l’activation des muscles antagonistes et une diminution de l’activation des muscles synergiques.
· Un facteur musculaire : Sa contribution se fait plus tard. Ils concernent l’hypertrophie du muscle où serait impliquée une hypertrophie des fibres (augmentation du diamètre), due à une augmentation du nombre et du diamètre des myofibrilles (augmentation des protéines contractiles) e une augmentation du tissu conjonctif qui rend le muscle plus résistant. Il y a aussi une hyperplasie (augmentation du nombre de fibres musculaires). Mais ce phénomène est faible.
C : Facteurs psychologiques :
Ils sont en rapport avec la motivation du sujet et leur influence est montrée par la stimulation du muscle. La force développée lors d’une contraction tétanique est 30% plus importante que celle développée lors d’une contraction normale.
Les facteurs psychologiques sont très liés à une modification du système nerveux central, ce qui modifie la fréquence des influx nerveux envoyés par les motoneurones.
5-5 : FATIGUE MUSCULAIRE :
5-5-1 : Définition :
C’est la capacité à développer une force programmée, donc une impossibilité pour le muscle de maintenir une contraction qui donne une force donnée. Quand on stimule le muscle par des stimulations linéaires et répétées, on assiste à une diminution de la force maximale et à une diminution de la vitesse de raccourcissement, dues au ralentissement important de la relaxation.
5-5-2 : Sites impliqués :
Ces différents sites peuvent avoir deux principales origines :
· Une origine centrale : L’information part du cortex, puis passe dans la moelle épinière et enfin dans le motoneurone.
· Une origine périphérique : L’information part de la plaque motrice (problème de transmission neuromusculaire). Il y a ensuite propagation de l’influx nerveux (sarcolème et tubule T) et libération de calcium (réticulum sarcoplasmique) ce qui joue sur les liaisons actine-myosine.
Au niveau de ces sites 4 sont le plus souvent impliqués :
· Les motoneurones α : La diminution de la fréquence d’émission des influx nerveux.
· La propagation de l’influx nerveux : Le sarcolème est plus excitable.
· Le réticulum sarcoplasmique : Diminution de calcium, augmentation de sa récupération.
· Les myofibriles : Diminution de la troponine et des mécanismes de contraction.
5-5-3 : Les différents mécanismes impliqués :
A : Activités des motoneurones :
Leur activité est ce qu’on appelle une fatigue d’origine centrale et montré par le fait que dans la fatigue, il y a une modification de l’électromyographie, qui correspond à l’activité électrique du muscle ( = Activation nerveuse du muscle). Quand on se fatigue, il y a une diminution de l’activité du muscle. Cette fatigue centrale est montrée par une diminution progressive de la force lors des contractions volontaires (force musculaire volontaire), alors que, quand on stimule le muscle directement, n retrouve la force initiale (ce qui n’est pas le cas quand le muscle est fatigué). Ca correspond à une modification (diminution) de la fréquence de décharge, qui implique deux mécanismes :
· Diminution de la stimulation des motoneurones par les centres nerveux supérieurs. Ce phénomène peut-être modifié (augmenter ou diminuer) par une émotion ou par tout autre stimulation (ex : Stimulation verbale). Pour les efforts de longue durée, on note une amplification de la stimulation centrale, qui est amplifiée par l’hypoglycémie.
· Une inhibition progressive des motoneurones par des influx afférents qui proviennent des fibres sensitives en provenance des muscles (exemple : Des informations qui viennent de nocicepteurs, qui vont être activées par des stimulations chimiques, mécaniques ou thermiques). C’est un mécanisme secondaire. La fatigue centrale est égale à 10% de la diminution de la force.
B : Propriétés du sarcolème :
Il peut y avoir des perturbations au niveau du sarcolème. Les propriétés qui ont été modifiées, consistent à une altération de la propagation du potentiel d’action, et de la transmission neuromusculaire. Et ceci n’expliquera pas une augmentation de la concentration de potassium dans le liquide extracellulaire : En dehors des activités du muscle. Le potassium joue donc un grand rôle dans le fonctionnement des fibres musculaires (détermine le potentiel de repos de la membrane, c’est-à-dire qu’il y a une différence entre l’intérieur et l’extérieur de la cellule musculaire). Il varie entre –70 mV (fibres lentes) et –80 mV (fibres rapides) ; Il est le résultat de la répartition des charges électriques de part et d’autre de la membrane (négatives à l’extérieur et positives à l’extérieur). Donc au repos, le milieu extracellulaire est riche en sodium et le milieu intracellulaire est riche en ions potassium. La concentration en sodium est plus grande à l’extérieur qu’à l’intérieur car la membrane est peu perméable au sodium et il existe une pompe Na+-K+ ATPase. Elle fait entrer deux potassiums dans la cellule et fait sortir en même temps trois sodiums.
L’intérieur est moins négatif car les ions négatifs sont bloqués à l’intérieur de la cellule : On les retrouve au niveau des acides aminés et des protéines phosphatées. La grande quantité des ces cations est plus grande à l’extérieur, car les ions sodiums restent bloqués à l’extérieur de la cellule et la pompe expulse les ions sodium. Par contre les potassiums sortent de la cellule, car la membrane est plus imperméable (50 à 100 fois) au potassium.
Un potentiel correspond à une dépolarisation, qui est due à une modification de la perméabilité de la membrane, c’est-à-dire une diminution de la différence des charges négatives entre l’intérieur et l’extérieur.
Au niveau de la plaque motrice, l’avancée d’un potentiel d’action provoque l’ouverture de canaux calciums, ce qui engendre l’entrée de calcium au niveau de cette terminaison pré-synaptique. Ceci induit la libération d’acétylcholine. Une fois libérer, il se fixe sur des récepteurs spécifiques sur le sarcolème ( = Membrane de la fibre musculaire). Ce récepteur spécifique (récepteur nicotique) est une protéine canale. Quand il se fixe, il y a ouverture du canal qui laisse passer les ions sodium et potassiums.
On observe donc le passage du sodium qui entre à l’intérieur, car il y en a beaucoup à l’extérieur. L’extérieur prend donc des ions sodiums et l’intérieur devient plus positif par rapport à l’extérieur. Le voltage passe donc de –70 mV à +30 mV. Quand le potentiel de membrane passe à 0, l’entrée sodium devient donc de plus en plus difficile. Cette entrée est donc transitoire. En réponse à cette augmentation de la charge positive à l’intérieur, il y a sortie par le même canal des potassiums. L’extérieur devient plus positif que l’intérieur. La différence de potentiel revient à –70 mV ( = Repolarisation). Mais on est dans les conditions inverses aux conditions de repos. Pour rétablir l’équilibre ionique de repos, les pompes font entrer de nouveau les potassiums dans la cellule et les sodiums vont ressortir. Cette pompe permet de maintenir une concentration faible en sodium et forte en potassium. Une modification de la concentration de ces deux cations va perturber le potentiel d’action, dans le sens où l’importance et la durée du potentiel est corrélé au courent de sodium et de potassium. S’il y a une faible concentration extracellulaire et une forte concentration intracellulaire en sodium, on note alors une diminution du flux entrant de cet ion ; Et inversement pour le potassium.
Aussi une accumulation de potassium induit une diminution du seuil d’excitation du sarcolème. Il faudra donc un potentiel plus grand. Au repos la concentration en potassium est égale à 5mmol. Si cette concentration passe à 12mmol, les fibres deviennent alors inexcitables. Cette augmentation de la concentration extracellulaire en potassium serait principalement impliquée dans la fatigue liée aux activités de grande intensité (fréquence de stimulation importante). Donc il y a accumulation de potassiums dans le liquide extracellulaire, qui activerait les pompes sodium-potassium ATPase. L’explication est due à un approvisionnement très faible en ATP du à une perturbation du métabolisme énergétique.
C : Accumulation de sous-produits
métaboliques :
L’énergie indispensable à la contraction des ponts actine - myosine vient de l’hydrolyse de l’ATP. Les différentes sources permettent un approvisionnement constant en ATP :
· Les sources immédiates d’énergie, sui conservent :
- L’ATP : Il y a des réserves d’ATP dans les fibres, mais qui sont faibles : 5 à 6 mmol.kg-1 de muscle (ce qui représente environ 10 à 20 mètres ou une contraction pendant 3 secondes).
- La phosphacrétine ( = Créatine phosphate), qui est une molécule, qui possède une liaison phosphate riche en énergie. La rupture permet de constituer très vite de l’ATP. La phosphocréatine est hydrolysée par une enzyme la créatine phosphate et l’énergie libérée permet de reconstituer de l’ATP et relient le phosphate inorganique à une molécule d’ADP. Mais les réserves sont également très faibles. : 25 mmol.KG-1 de muscle (exercice intense pendant 20 à 30 secondes).
· Les systèmes énergétiques :
- Le métabolisme glycoltique, qui augmente dans les première seconde de l’exercice et qui correspond à la glycolyse c’est-à-dire la dégradation du glucose ou glycogène en acide pyruvique. La glycolyse comprend 10 réactions et se fait dans le sarcoplasme. Elle produit peu d’ATP, à savoir 2 par mole de glucose ou 3 par molécule de glycogène. La réaction ne nécessite pas d’oxygène et sans oxygène l’acide pyruvique est réduit en acide lactique.
- Le métabolisme oxydatif : Il est sollicité après quelques minutes d’exercice, quand l’apport d’oxygène augmente. Il dégrade les substrats en présence d’oxygène dans les mitochondries. Il fait intervenir le cycle de Krebs, la béta-oxydation et la chaîne de transport des électrons.
Le cycle de Krebs est l’oxydation de l’acétyl-CoA en Carbonne et en hydrogène. L’acétyl-CoA provient soit de la béta-oxydation ( = Dégradation des acides aminés en acétyl-CoA). Le carbone se lie à de l’oxygène pour former du CO2 qui est éliminer pendant l’expiration. Les hydrogènes formés seront pris en charge par des Co-enzymes qui vont les transporter vers la chaîne de transport des mitochondries. Les hydrogènes formés dans le cycle de Krebs passent dans une série d’oxydo-réduction et sont divisés en protons et en électrons et c’est ce transport des électrons qui fournit de l’énergie qui permet la phosphorylation de l’ATP en ADP.Ce processus nécessite de l’oxygène car c’est le transporteur final de l’électron. Le rendement énergétique est important. Il y a formation de 38 molécules d’ATP par molécule de glucose et plus de 100 molécules d’ATP par acides gras.
La fatigue pendant un exercice de courte durée impliquerait une accumulation de sous-produits métaboliques et notamment le phosphate inorganique et les ions hydrogènes.
Le phosphate inorganique : Une augmentation de la concentration en phosphate inorganique lors de la fatigue, qui est due à la dégradation de l’ATP. Cela diminue beaucoup la force développée par un muscle et la vitesse de contraction (mais de moindre importance). Une augmentation de la concentration en phosphate inorganique altère les liaisons actine – myosine et cette altération est plus importante dans les fibres rapides que dans les fibres lentes. L’hypothèse est donc que le phosphate inorganique joue au niveau de la myosine et diminue l’activité de l’ATPase. De plus une augmentation importante de la concentration en phosphate inorganique est capable de réduire la quantité d’énergie lors de l’hydrolyse de l’ATP. D’autre part cette augmentation peut être impliquée dans l’augmentation de la relaxation en perturbant le fonctionnement des pompes calcium.
Les ions hydrogènes : Une entrée des ions hydrogènes est due à l’accumulation d’acide lactique lors d’une mise en jeu importante du métabolisme glycolitique. Il s’accumule à l’intérieur des fibres musculaires. S’il n’est pas éliminé rapidement, il se divise en ions lactate et hydrogènes. Il y a donc une augmentation des ions hydrogènes et une diminution du pH, qui conduit à l’acidose musculaire. Mais les ions hydrogènes sont diffusés en dehors des cellules musculaires et passent dans le sang (idem pour le lactate). Il y a alors diminution du pH sanguin.
Il existe au niveau musculaire et sanguin, des molécules tampons, qui neutralisent l’action des ions hydrogènes. S’il n’y avait pas de tampon le pH diminuerait de 1.5. Lors d’une activité très intense le pH passe de 7.1 à au maximum 6.4, mais une faible diminution de pH entraîne des perturbations dans la production d’énergie et la contraction musculaire.
Au niveau du métabolisme énergétique, la diminution du pH provoque donc une diminution de la resynthèse de l’ATP. Un pH de 6.9 inhibe la glycolyse et il y a alors rapidement diminution de la quantité d’ATP.
Au niveau de la concentration musculaire, on remarque une diminution de la force et de la vitesse de contraction : Comment ?
· En diminuant la sensibilité de la troponine au calcium. Celle-ci le capte donc moins et pour une augmentation de calcium, la force produite sera plus faible. La sensibilité est diminuée de moitié quand le pH diminue de 0.5.
· Cette diminution joue un rôle direct au niveau des liaisons actine – myosine : Il y a en effet diminution du nombre et de la force générée par chacune des liaisons.
· La diminution de la fréquence du détachement des liaisons actine – myosine : Il y a donc diminution de la vitesse de contraction.
Aussi il y a une différence entre les fibres rapides et les fibres lentes, car les fibres rapides ont un métabolisme glucidique préférentiellement : Il y a donc plus d’acide lactique ainsi qu’une diminution du pH plus importante dans les fibres rapides.
La diminution de pH est aussi responsable de la diminution de la relaxation, car elle perturbe le fonctionnement des pompes calcium ATPase du réticulum sarcoplasmique. La diminution du pH serait la principale cause de la fatigue lors d’un exercice bref et intense. Il faut entre 30 et 35 minutes pour rétablir le pH après ce type d’effort.
D : Epuisement des substrats
énergétiques :
Cela concerne les exercices de longue durée et qui concerne l’épuisement des réserves de glucide. Il y a épuisement des stocks de glycogènes qui concernent :
· L’épuisement du glycogène lipidique.
· L’épuisement du glycogène musculaire.
Ceci induit une hypoglycémie et qui est susceptible de perturber la performance des exercices de plus de 30 minutes
Pourquoi ?
· Le glycogène est un substrat essentiel. Sans glycogène, il n’y a pas de synthèse d’ATP.
· Il est indispensable à la libération de calcium par le réticulum sarcoplasmique.
· L’hypoglycémie amène à des effets au niveau de la fatigue centrale.
VI – CONTROLE NERVEUX DU
MOUVEMENT :
6-1 : GENERALITES :

Le contrôle nerveux du mouvement a trois principales fonctions :
· Fonction sensorielle.
· Fonction intégrative : Permet d’analyser les données sensorielles reçues, d’en stocker des aspects.
· Fonction motrice : Peut réagir aux informations reçues en provoquant des contractions musculaires ou des sécrétions glandulaires par le système nerveux autonome.
Le système nerveux est composé de neurones, de cellules de soutien (cellules gliales : Cellules de SCHWANN).
Un ganglion est un regroupement de corps cellulaires.
6-2 : SYSTEME NERVEUX CENTRAL :
Le système nerveux central comprend 1012 neurones englobe la substance blanche ( fibres myélinisées)et la substance grise (corps cellulaires des neurones). Il est constitué de différentes parties.
6-2-1 : L’encéphale :
L’encéphale pèse environ 1.5 kg et est divisé en 4 régions :
· Le cerveau, qui est constitué de deux hémisphères et enveloppé par le cortex cérébral. Le cortex cérébral est le siège de la pensée et de l’intellect. On lui donne aussi le nom de substance grise, car il est composé uniquement de neurones. C’est l’écorce du cerveau.
C’est la partie consciente du cerveau, qui permet de penser, d’avoir conscience des informations sensorielles, de contrôler les mouvements. Il est divisé en ères de types sensitives (reçoivent et interprètent les influx sensitifs), motrices (contrôlent l’acte moteur, c’est-à-dire l’activité musculaire) et les aires d’association.
Le cerveau est lui-même composé de 4 lobes, qui recueillent chacun une partie des informations transmises par les récepteurs :
- Le lobe frontal : Il a une fonction intellectuelle, contrôle l’odorat et le contrôle moteur.
- Le lobe temporal, qui intervient dans les perceptions auditives.
- Le lobe pariétal, qui reçoit toutes les informations sensitives.
- Le lobe occipital : Il gère les informations visuelles.
· Le diencéphale composé du thalamus et de l’hypothalamus. Le thalamus est un centre d’intégration très important : Toutes les fonctions sensorielles sauf l’odorat y sont relayées. Il relaye aussi les influx moteurs, qui vont du cortex cérébral jusqu’à la moelle épinière.
L’hypothalamus est responsable du maintien de l’homéostasie, car il régule tous les processus qui vont agir sur le milieu interne. Il reçoit des signaux hormonaux et des influx sensitifs venant des viscères. Il contrôle l’activité du système nerveux autonome (pression artérielle, fréquence cardiaque). Il est le lien principal entre le système endocrinien et le système nerveux, car il sécrète des hormones qui contrôlent l’hypophyse. Il contrôle la température, la soif et les cycles éveil - sommeil.
· Le tronc cérébral : Il est composé de trois parties :
- Les pédoncules cérébraux.
- Le bulbe rachidien.
- La protubérance = Le pont.
Il assure la fonction entre le cerveau et la moelle épinière. Par conséquent tous les nerfs (moteurs ou sensitifs) traversent le tronc cérébral. On y retrouve la plupart des centres régulateurs du système nerveux autonome (ex : Le centre cardiovasculaire, le centre respiratoire).
On y retrouve aussi un ensemble de neurones : La formation réticulée, qui est en interrelation permanente avec toutes les autres régions du système nerveux central.
· Le cervelet : Il est en relation avec le cerveau et joue un rôle central dans le rôle du mouvement (Rôle essentiel dans la conduite motrice, mais aussi dans la posture et l’équilibre)
6-2-2 : La moelle épinière :
Elle se situe juste au-dessous de l’encéphale. C’est le prolongement du bulbe rachidien et donc de l’encéphale. Elle est dans la colonne vertébrale du bulbe rachidien jusqu’à la deuxième vertèbre lombaire (longueur = 45 cm et diamètre = 2 cm). La moelle épinière est le lieu de passage de toutes les fibres motrices ou sensitives qui transitent entre l’encéphale et la périphérie de l’organisme.
Elle est aussi un site d’intégration de ces influx nerveux. Elle contient de la substance grise (forme de papillon) entourée de substance blanche. La substance grise est subdivisée en région : Les cornes (corne antérieure, coté ventral ; corne postérieure, coté dorsal). Entre les deux se trouvent la corne latérale. Il y arrive des neurones moteurs ou d’association. La substance grise reçoit et interprète l’information sensitive (qui entre) et l’information matrice (sui sort). La substance blanche contient les axones, qui sont myélinisés : Ce sont soit des neurones moteurs, soit des neurones sensitifs.
6-3 : LES SYSTEME NERVEUX
PERIPHERIQUE :
Il correspond à 43 paires de nerfs :
· 12 paires de nerfs crâniens.
· 31 paires de nerfs rachidiens, qui viennent de la moelle épinière.
10 nerfs crâniens proviennent du tronc cérébral. Certains ne sont que sensitifs, d’autres que moteurs et les derniers sont mixtes.
Dans les nerfs rachidiens, 8 paires sont des nerfs cervicaux, 12 des nerfs dorsaux, 5 des nerfs lombaires, 5 des nerfs sacrés et 1 nerf coccidien. Ils viennent des régions spécialisées de la peau : Les dermatoses. Ils sont reliés au système nerveux central, c’est-à-dire la moelle épinière par leur racine postérieure et une racine antérieure. Ils sont donc mixtes (une voix motrice et une voix sensitive).
La racine postérieure contient des fibres sensitives. On y remarque aussi un renflement, qui est le ganglion rachidien, qui contient le corps cellulaire de neurone sensitif. La racine dorsale contient les axones des neurones moteurs. Elle conduit les informations jusqu’à l’effecteur. (Pour nous c’est le muscle). Le corps cellulaire est directement dans la substance grise et innerve directement la muscle squelettique. L’axone peut donc être très long.
6-3-1 : Voix sensitives :
Les voix sensitives amènent des informations en provenance de tout l’organisme (vaisseaux sanguins, organes internes, organes des sens, de la peau, des muscles et des tendons)
Il y a 5 grands types de récepteurs :
· Les mécanorécepteurs, sensibles aux stimulations mécaniques.
· Les thermorécepteurs, sensibles aux variations de la lumière.
· Les nocicepteurs, sensibles aux stimulations de douleurs.
· Les photorécepteurs, sensibles aux variations de la lumière.
· Les chémorécepteurs ( = Chimiorécepteurs), sensible aux modifications chimiques de l’organisme.
Ils se terminent dans le système nerveux central et informent donc les modifications qui se passent dans le corps.
6-3-2 : Les voix motrices :
Les informations sensorielles sont traitées au niveau de centres nerveux supérieurs et la réponse appropriée est envoyée aux effecteurs.
6-4 : INTEGRATION
SENSORI-MOTRICE :
Comment fonctionnent les centres sensori-moteurs pour répondre ?
6-4-1 : Informations sensitives :
Elles sont transmises par l’intermédiaire des nerfs sensitifs à la moelle épinière. Chaque axone qui y pénètre s’y divise en plusieurs branches :
· Une branche segmentaire, qui reste au niveau de la moelle épinière.
· Une branche montante.
· Une branche descendante.
L’information sensitive va diffuser rapidement vers la moelle épinière et l’encéphale. Le mode d’intégration varie selon le niveau où arrive cette information sensitive :
· Si elle atteint la moelle épinière, la réponse est un simple réflexe (rapide et inconscient).
· Si elle atteint l’encéphale, les informations peuvent s’y terminer dans différentes régions :
- Si elles se terminent dans le tronc cérébral inférieur ou le cervelet, cela entraîne une réponse motrice subconsciente. Ca implique la coordination de différents muscles. On appelle cela la proprioception (consciente).
- Si elles se terminent dans le thalamus (plus haut), l’information atteint notre conscience et provoque différentes sensations.
- Si elles se terminent dans le cortex cérébral : On peut percevoir le lieu d’origine de ces informations.
6-4-2 : Le contrôle moteur :
Quel que soit le niveau atteint par l’information sensitive, celle-ci déclenche la réponse d’un motoneurone, qui va stimuler les muscles squelettiques. Au fur et à mesure que le niveau de contrôle augmente, les mouvements vont devenir de plus en plus complexes. Le mouvement le plus simple étant un réflexe.
Au niveau de ces motoneurones, il en existe deux types :
· Les motoneurones α, qui innervent les muscles squelettiques. Ce sont des cellules de grande taille (diamètre entre 60 et 80 µm et des axones de diamètre proche de 9µm). Ils sont impliqués dans la commande motrice.
· Les motoneurones γ, qui innervent un type de mécanorécepteur musculaire : Les fuseaux neuromusculaires. Ils sont plus petits et deux fois moins nombreux. L’axone a un faible diamètre (5 à 7 µm)
· Les interneurones : Ils établissent des connexions locales avec les motoneurones où envoient des informations vers les centres supérieurs.
6-4-3 : Activité réflexe :
C’est une réponse préprogrammée. Il s’agit d’un réflexe qui intervient dans le contrôle de la contraction musculaire ce qui nécessite l’intervention du motoneurone α et un système nerveux sur l’état du muscle (état sur la longueur ou sur la force). Cela concerne deux types de récepteurs :
· Les mécanorécepteurs sensitifs musculaires.
· Les organes tendineux de Golgi.
A : Fuseaux neuromusculaires :
Ils sont à l’intérieur des fibres squelettique. Leur diamètre est de 100µm et leur longueur de 10mm. Ils sont constitués de 3 à 12 fibres musculaires striées, mais qui sont modifiées dans le sens où la région centrale des fibres ne contient pas d’actine ou de myosine. Elles sont appelées intrafusales. L’extrémité du fuseau est rattachée au tissu conjonctif du muscle. Les extrémités des fibres du fuseau sont innervées par des motoneurones γ. La partie centrale des fibres du fuseau constitue les récepteurs sensoriels sensibles à l’étirement. Ils sont stimulés quand le récepteur s’allonge. La terminaison centrale est innervée par deux types de terminaisons sensitives :
· Les terminaisons primaires : Ia. Elles conduisent l’influx sensitif à une très grande vitesse (70 à 120 ms). Elles sont sensibles à la longueur et à la vitesse de changement de longueur du muscle. Ceci donne une réponse très dynamique, qui dépend de la vitesse d’étirement du muscle, puis qui donne une réponse statique pendant tout le temps de l’étirement.
· Les fibres secondaires : II, qui sont plus petite (vitesse de contraction plus faible) et qui ne sont sensibles qu’à l’étirement du muscle.
Le choix se fait donc uniquement en fonction de la vitesse de l’étirement : Si elle est rapide, seules les terminaisons primaires seront stimulées (réponses intenses et dynamiques). Si la vitesse est lente, les primaires et les secondaires seront stimulées.
Par ces deux types de terminaison le système nerveux sera impliqué dans un réflexe d’étirement : Le réflexe myotatique. Si le muscle est étiré, il en résulte une contraction réflexe, qui met en jeu un circuit neuronal, qui comprend :
· Le fuseau neuromuscualire.
· Les fibres sensitives primaires, qui apportent l’information au système nerveux.
· Le motoneurone α et son axone qui est stimulée par les fibres sensitives.
· Le muscle d’où provient la fibre Ia du fuseau.
Ce réflexe contient deux composantes :
· Une composante phasique : Le réflexe d’étirement phasique est déclenché par l’activation des terminaisons primaires lors d’un étirement très brutal. Ceci induit une contraction réflexe du muscle d’où provient le signal. Un tel réflexe est court (30 ms), car les fibres Ia arrivent à la moelle épinière par la corne dorsale et finissent sur le muscle. Les fibres Ia émettent une collatérale, qui se termine sur un interneurone, qui va inhiber le motoneurone α qui innerve le muscle antagoniste.
· Une composante tonique.
Le réflexe myotatique a deux composantes: Une composante tonique et une composante phasique. La composante phasique se produit après un étirement violent et dur. La composante tonique est arrive suite à un étirement plus doux. Dans cette composante, les deux types de terminaisons sont stimulés. Le message est statique et continu. Les terminaisons sensitives de type deux se terminent sur des interneurones, qui vont stimuler le motoneurone alpha, qui innerve le muscle et un deuxième interneurone qui inhibe le motoneurone qui innerve le muscle: Il s'agit du réflexe polysynaptique. Ce réflexe d'étirement tonique joue un rôle important dans le maintien du tonus et de la posture.
Les fuseaux jouent un rôle dans la contraction normale du muscle. Ils sont innervés par des motoneurones gamma, qui innervent les extrémités du fuseau neuromusculaire.
Quand il y a contraction involontaire, il y a activation des deux types de motoneurones (coactivation des motoneurones alpha et gamma). Si les motoneurones gamma sont activés, il y a contraction des fibres aux extrémités ce qui provoque la contraction de la partie centrale. Les fibres sensitives Ia et II sont activées et vont elles-mêmes activées le motoneurone gamma. Les impulsions nerveuses apportées par les fibres sensitives peuvent monter plus haut et atteindre le système nerveux central. Le cerveau connaît alors la longueur du muscle. Ces informations permettent de maintenir le tonus musculaire.
B: Les
organes tendineux de Golgi:
C'est un récepteur sensoriel constitué de collagène et entouré d'une capsule conjonctive. Son diamètre est de 100 micromètres et une longueur de 1mm. Il est situé aux extrémités proximales des tendons à la jonction tendons-muscles. Une des extrémités est insérée sur le tendon et l'autre est reliée à des fibres musculaires (entre 5 et 25 fibres musculaires) appartenant à plusieurs unités motrices différentes. Réciproquement une unité motrice peut activer de 4 à 6 organes tendineux différents.
L'organe tendineux est sensible à la force développée par les fibres sur lesquelles il est fixé. Il est capable de produire une réponse statique, proportionnelle à la force, mais aussi une réponse dynamique quand il y a une variation brutale de la force.
Quand il est stimulé, il transmet ses informations à la moelle épinière par des fibres sensitives de type Ib. Cette fibre Ib active un interneurone, qui inhibe le motoneurone alpha d'où provient l'organe tendineux. Il y a également inhibition du motoneurone alpha du muscle antagoniste. On appelle cela le réflexe myotatique inversé.
S'il y a une
diminution des influx au niveau des fibres Ib, il y a une augmentation de
l'excitabilité du motoneurone alpha et donc une augmentation de la force
développée par le muscle. Quand le muscle développe une force importante, il y
a stimulation des organes tendineux de Golgi ce qui inhibe le motoneurone
alpha. Le but est la protection du muscle contre les effets néfastes d'une
contraction trop grande et de contrôler la force musculaire : Si elle est
élevée, l'organe tendineux fait en sorte de la diminuer et inversement si elle
est faible.
6-4-4:
Les centres nerveux supérieurs:
A: Les
cortex moteur:
Il est dans le lobe frontal et est responsable du contrôle des mouvements fins et précis. Il décide du mouvement à réaliser par les muscles. Il comprend le cortex moteur primitif (ou primaire): L'aire 4. Il assure la commande motrice et est donc impliqué dans l'exécution. A coté il y a le cortex commandant un ou plusieurs muscles dans des organes précieux du corps. Ces régions qui ont besoins de précision ont une grande représentation au niveau du cortex. Par exemple, les muscles de la main occupent plus de la moitié du cortex moteur
Ce contrôle conscient du mouvement des muscles squelettique est assuré par un type de motoneurone, qu'on appelle les cellules pyramidales. Leurs corps cellulaires se trouvent dans le cortex moteur primitif et leurs axones vont descendre dans la moelle épinière. C'est ce qu'on appelle le cortex pyramidal ou corticospinal, qui constitue la partie la plus importante des voix motrices descendantes. On les appelle voix pyramidales, car elle a une forme de pyramide inversée. Les neurones dont le corps cellulaire se trouve dans le cortex commande des muscles qui sont dans l'autre moitié du corps. L'hémisphère gauche commande les muscles de droite et inversement.
C'est un lieu de
convergence. Il reçoit des informations du cerveau mais aussi du thalamus, qui
reçoit lui-même des informations du cervelet. Il reçoit donc des informations
de l'ensemble du corps mais aussi de l'extérieur (informations visuelles et
auditives). Ca lui permet d'effectuer une commande motrice appropriée. Ces voix
assurent l'essentiel du contrôle de l'activité volontaire. Ce contrôle est ainsi
sous la dépendance d'autres structures.
B: Les
noyaux gris centraux:
Ce sont des structures sous corticales (sous le cortex cérébral). Il a un rôle très important dans:
· L'initiation des mouvements automatiques et répétés.
· Le contrôle des mouvements complexes et semi-volontaires.
· Implication dans le tonus musculaire et le maintien de la posture.
Ils
reçoivent des informations sensorielles et sont informés de ce qui se passe en
périphérie. Ils contrôlent les actes moteurs en association avec le cortex cérébral,
mais peuvent aussi agir sans le cortex.
C: Le
cervelet:
Il
ne dispose pas de la capacité à déclencher un mouvement par lui-même et est
indispensable à la réalisation d'un mouvement précis et harmonieux grâce à la
mise en jeu de plusieurs muscles. Il est placé en dérivation par rapport aux
voix motrices et perçoit en permanence des informations venant du cerveau, mais
aussi des informations sensorielles de la périphérie (visuelles, vestibulaires,
…). Il compare les informations au programme moteur en cours et apporte les
modifications nécessaires.
VII – FORMATION
ET DEVELOPPEMET DU MUSCLE SUQELETIQUE:
7-1:
INTRODUCTION:
Au cours du développement, il y a mise en place du premier feuillet
embryonnaire (ou cellulaire). C'est ce qu'on appelle la gastrulation. Ensuite
il y a la mise en place des organes: L'organogénèse. Chez l'homme tout part de
la fécondation, qui donne un œuf. Dans la première semaine, il se sépare et se
divise pour donner un blastocyte. Il se divise en blastomères (Première heure:
8 blastomères). La deuxième semaine après la fécondation, la masse de cellules
s'incruste dans la muqueuse de l'utérus partir de là commence à se former les
trois feuillets embryonnaires (C'est ce qu'on appelle la gastrulation). Il
s'agit de:
· L'ectoderme.
· L'endoderme.
· Le mésoderme.
Tous les
tissus et tous les organes du corps sont issus des organes embryonnaires.
7-2: ORIGINE
EMBRYONNAIRE DU MUSCLE SQUELETTIQUE:


7-3 :
FORMATION DES FIBRES MUSCULAIRES :
7-3-1:
Les différentes étapes:
Cf schéma.
A la naissance, la majorité des fibres sont formée. Certains myoblastes ne fusionnent pas et resteraient entre le sarcolème et une deuxième membrane: La lame basale. Ca représente environ 3 à 4% des noyaux de la fibre. On les appelle les cellules satellites qui servent à réparer le muscle. Elles sont au repos: Elles ne se divisent plus, mais peuvent être activées et reprendre leur cycle et fusionner avec la fibre (exemple: Entraînement).
7-3-2:
Facteurs myogéniques:
Ce sont des facteurs de transcription: Ce sont des protéines capables de se fixer sur l'ADN et stimule la formation de l'ARNm. Elles sont dans le noyau et on n les trouve que dans le muscle squelettique. Ces facteurs sont capables d'agir sur des protéines qui codent que pour e muscle squelettique. Elles peuvent convertir une cellule quelconque en cellule musculaire. Les facteurs sont au nombre de 4:
·
MYO D.
·
MYOGENINE.
·
MYF 5.
·
MRF 4
(MYF 6).
Ces facteurs agissent sous forme de complexe (association de deux facteurs myogéniques ou d'un facteur myogénique et d'une autre protéine). La fixation se fait sur l'ADN au niveau des boites E (succession particulière de bases).
Leur rôle est de:
· MYO D et MYF 5: Détermination: Passage entre les cellules du mésoderme aux myoblastes. Il faut les inactiver pour qu'il y ait fonction de myoblastes.
· Myogénine et MRF 4: Différentiation. La myogénine est indispensable à la fusion des myoblastes. La MRF 4 joue un rôle dans la différentiation tardive.
· MRF 4: Maturation en fibres musculaires.
7-4: APPARITION
ET MODULATION DE LA DIVERSITE MUSCULAIRE:
Comment peut-il exister différents types de fibres?
Les deux mécanismes impliqués seraient :
· La prédétermination dans les myoblastes.
· Une modulation extérieure de la maturation du muscle.
7-4-1:
Prédétermination:
Au niveau des myoblastes, il faut savoir que le muscle est formé par deux
générations successives de myotubes:
· Il y aurait donc une première migration qui donne les myoblastes primaires qui fusionnent pour donner les myotubes primaires.
· Il y aurait ensuite une seconde migration qui donne les myoblastes secondaires et ils fusionnent pour former des myotubes secondaires. Mais certains myoblastes secondaires fusionnent avec des myotubes primaires.
· Il y aurait une troisième migration, qui serait les cellules satellites.
La mise en
évidence de ce décalage a montré qu'un type de myoblaste donne un type de
fibres. Les myotubes primaires donnent les fibres I et les myotubes secondaires
donnent les fibres II.
7-4-2:
Modulation extrinsèque de la maturation du muscle:
Les
myoblastes secondaires et les cellules satellites sont plus sensibles au milieu
extérieur et sont donc plus modulables.
A: Le rôle
du nerf:
Les expériences d'innervation croisée, qui consistent à greffer le nerf d'une fibre rapide sur une fibre lente. On voit que le type de la fibre change. Mais la transformation n'est jamais complète. On s'est aperçu que si on dénnervait des fibres, il y avait dégénérescence plus grande chez les fibres lentes, qui deviennent des fibres rapides. Mais ce n'est pas le cas dans l'autre sens; L'innervation joue donc un rôle important chez est fibres lentes (Au niveau du phénotype lent). Le phénotype rapide est dit phénotype par défaut.
L'innervation est essentielle pour le maintien des fibres lentes. Cela s'explique aussi par le fait que les nerfs produisent des facteurs, appelés facteurs trophiques.
B: Rôle
des hormones thyroïdiennes:
Elle joue un rôle
exclusivement sur les fibres rapides. Si on supprime la thyroïde, il y a
diminution du pourcentage des fibres IIb mais pas des fibres I. Par conséquent
elles sont indispensables à la formation de MHC IIb, et donc d'un phénotype
rapide. Les fibres IIa sont moins touchées.
C: Rôle
des facteurs de croissance:
Ils
sont capables de contrôler la différence et la prolifération des myoblastes.
Parmi eux:
· IGF 1: Il peut être produit par le foie via des hormones de croissance, qui peut être produit au niveau du muscle. Il est capable de stimuler la prolifération et la différentiation.
· FGF, qui peuvent augmenter la prolifération, mais qui inhibe la différentiation.
· HGF: Idem.
· TGF béta: Il stimule et inhibe selon le cas la prolifération et la différentiation.